 2015-05-26
2015-05-26 10979
10979«Наиболее выдающаяся черта в жизни растения заключается в том, что оно растет: на это указывает самое название его».
К. А. Тимирязев
В предыдущей главе были рассмотрены процессы, связанные с превращением питательных веществ в вещества и структуры самой клетки и этапы ее развития. Все эти процессы, несомненно, относятся к явлениям роста и развития растительного организма в целом. Однако целесообразнее было обсудить внутриклеточные процессы предварительно, так как ведущим фактором роста многоклеточного организма является взаимодействие клеток с участием межклеточных систем регуляции и под контролем доминирующих центров.
Несколько слов о терминах, применяемых при изучении роста и развития растений.
Онтогенезом (от греч. «on», род. падеж «ontos» — существо, лат. genesis — происхождение, процесс образования) называют индивидуальное развитие организма от зиготы (или вегетативного зачатка) до естественной смерти. В ходе онтогенеза реализуется наследственная информация организма (генотип) в конкретных условиях окружающей среды, в результате чего формируется фенотип, т. е. совокупность всех признаков и свойств данного индивидуального организма.
Развитие — это качественные изменения в структуре и функциональной активности растения и его частей (органов, тканей и клеток) в процессе онтогенеза. Возникновение качественных различий между клетками, тканями и органами получило название дифференцировки. В понятие «развитие» входят также и возрастные изменения.
Рост — необратимое увеличение размеров и массы клетки, органа или всего организма, связанное с новообразованием элементов их структур. Понятие «рост» отражает количественные изменения, сопровождающие развитие организма или его частей.
Если считать, что термины «развитие растений» и «онтогенез» тождественны, то соподчинение всех этих понятий можно представить в виде следующей схемы:

1 См: М. X. Чайлахян и др. Терминология роста и развития высших растений. М., 1982.
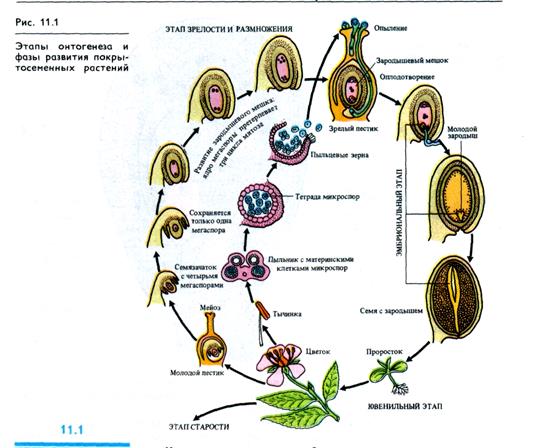
Развитие высших растений подразделяют на четыре этапа:
 1) эмбриональный, 2) ювенильный (молодость), 3) репродуктивный (зрелость), 4) старость (рис. 11.1).
1) эмбриональный, 2) ювенильный (молодость), 3) репродуктивный (зрелость), 4) старость (рис. 11.1).
Эмбриональный этап онтогенеза семенных растений охватывает развитие зародыша от зиготы до созревания семени включительно (см. рис. 11.1).
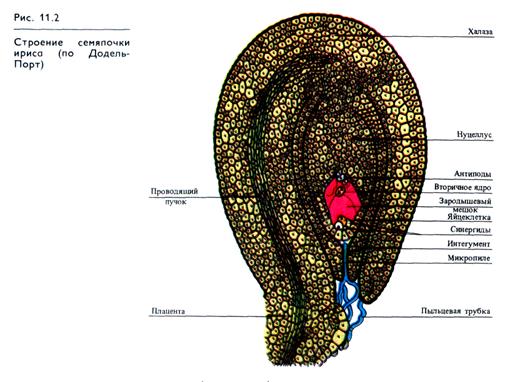
 Зигота у покрытосеменных образуется в результате слияния спермия пыльцевой трубки (мужской геметофит) с яйцеклеткой зародышевого мешка (женский гаметофит). В зародышевом мешке у покрытосеменных происходит открытое С. Г. Нава-шиным двойное оплодотворение, состоящее в слиянии второго спермия пыльцевой трубки со вторичным (диплоидным) ядром центральной клетки зародышевого мешка, что в дальнейшем приводит к образованию эндосперма. Зародышевый мешок (рис. 11.2) находится в нуцеллусе, окруженном интегументом (от лат. integumentum — покров). Нуцеллус используется для питания зародыша и реже превращается в запасающую ткань — перисперм.
Зигота у покрытосеменных образуется в результате слияния спермия пыльцевой трубки (мужской геметофит) с яйцеклеткой зародышевого мешка (женский гаметофит). В зародышевом мешке у покрытосеменных происходит открытое С. Г. Нава-шиным двойное оплодотворение, состоящее в слиянии второго спермия пыльцевой трубки со вторичным (диплоидным) ядром центральной клетки зародышевого мешка, что в дальнейшем приводит к образованию эндосперма. Зародышевый мешок (рис. 11.2) находится в нуцеллусе, окруженном интегументом (от лат. integumentum — покров). Нуцеллус используется для питания зародыша и реже превращается в запасающую ткань — перисперм.
Яйцеклетка в зародышевом мешке уже поляризована, т. е. две ее половины, обращенные к микропиле (пыльцевходу) и к халазе (основанию), имеют разное строение: ядро сдвинуто к халазальному полюсу, а в микропилярной половине расположена крупная вакуоль.
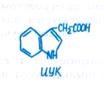 Зародыши проходят ряд последовательных фаз развития. Для большинства двудольных это проэмбрио, глобулярная, сердцевидная, торпедовидная (торпедо) и созревания (рис. 11.3).
Зародыши проходят ряд последовательных фаз развития. Для большинства двудольных это проэмбрио, глобулярная, сердцевидная, торпедовидная (торпедо) и созревания (рис. 11.3).
После оплодотворения зигота некоторое время (от нескольких часов до нескольких суток) находится в латентном состоянии. За это время в ней возрастает синтез РНК, она увеличивается в объеме. Триплоидное ядро центральной клетки зародышего мешка начинает делиться первым, образуя эндосперм. Для развития эндосперма нужны ИУК и цитокинин, которые поступают из нуцеллуса и из плаценты (место прикрепления семяпочки к стенке завязи).
При первом делении зиготы, плоскость которого перпендикулярна к оси ее поляризации, дочерняя клетка, обращенная к микропиле, выглядит более крупной и при дальнейшем делении образует однорядную нить клеток, называемую суспензором (подвеском). Функции суспензора: 1) удлиняясь, вдвигать развивающийся зародыш в ткань эндосперма, 2) служить гаусторией, поглощающей вещества из нуцеллуса и интегумента и передающей их зародышу, 3) синтезировать фитогормоны. Часть клеток суспензора, примыкающая к зародышу, у некоторых видов позднее входит в его состав в качестве апикаль ной меристемы и корневого чехлика зародышевого корня.
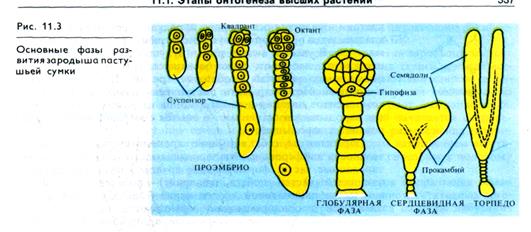
 Две синергиды, примыкающие к яйцеклетке, выполняют функцию гаусторий, поглощая вещества из нуцеллуса. Аналогичную функцию гаусторий осуществляют и клетки-антиподы, находящиеся рядом с эндоспермом.
Две синергиды, примыкающие к яйцеклетке, выполняют функцию гаусторий, поглощая вещества из нуцеллуса. Аналогичную функцию гаусторий осуществляют и клетки-антиподы, находящиеся рядом с эндоспермом.
Вторая клетка у двухклеточного зародыша, обращенная к эндосперму и халазальному полюсу, дважды делится в продольном по отношению к первичной оси поляризации направлении, образуя квадрант (рис. 11.3). Затем каждая из четырех клеток будущего зародыша у многих двудольных делится поперек, в результате чего формируется октант. Четыре дистальные (периферические) клетки октанта при дальнейшем делении сформируют семядоли и апекс побега, а четыре проксимальные — гипокотиль и базальную часть корня. Однако на этой фазе развития зародыша еще нет видимой дифференциации клеток, и зародыш, состоящий последовательно из одной, двух, четырех и восьми клеток, называют проэмбрио. По мере развития зародыша концентрация ИУК в клетках постепенно возрастает, происходит постоянный приток цитоки-нина. Последняя клетка суспензора, примыкающая к октанту, становится гипофйзой — инициальной клеткой корневого полюса и морфологически выделяется (рис. 11.3).
 Все восемь клеток проэмбрио делятся периклинально (т. е. плоскость их деления параллельна поверхности октанта), по-видимому, из-за распределения сил механического давления, направленного изнутри к поверхности. Эта фаза развития называется глобулярной. Наружные клетки в дальнейшем сформируют протодерму, а внутренние дадут начало первичной коре и центральному цилиндру. На этой фазе особенно необходим цитокинин. Фитогормоны поступают главным образом из развивающегося эндосперма, который играет ведущую роль в развитии зародыша.
Все восемь клеток проэмбрио делятся периклинально (т. е. плоскость их деления параллельна поверхности октанта), по-видимому, из-за распределения сил механического давления, направленного изнутри к поверхности. Эта фаза развития называется глобулярной. Наружные клетки в дальнейшем сформируют протодерму, а внутренние дадут начало первичной коре и центральному цилиндру. На этой фазе особенно необходим цитокинин. Фитогормоны поступают главным образом из развивающегося эндосперма, который играет ведущую роль в развитии зародыша.
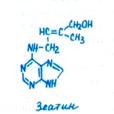
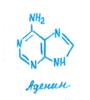
Следующая фаза развития зародыша получила название сердцевидной. В морфологически верхней части глобулярного зародыша наблюдается интенсивное билатеральное делениеклеток (появляется ось второго порядка), в результате чего закладываются примордии двух симметрично расположенных семядолей. В участке между ними (будущий апекс побега) деление клеток, наоборот, резко замедляется. В этот период для нормального развития необходим приток ИУК, цитокинина и аденина. Необходимость поступления извне тех или иных компонентов питания и фитогормонов на разных фазах развития зародыша устанавливают в опытах с выращиванием изолированных зародышей.
 Обращает на себя внимание хронологическая последовательность дифференцировки зародыша на отдельные специализированные части: формирование группы клеток зародыша — инициалей частей побега (квадрант) -»появление гипофизы в качестве инициальной клетки корня -> закладка примордиев семядолей с одновременным торможением делений клеток в участке между ними и появление прокамбия. Усложнение внутренних корреляционных взаимоотношений между клетками разных участков зародыша, по-видимому, указывает на становление собственной гормональной системы будущего организма.
Обращает на себя внимание хронологическая последовательность дифференцировки зародыша на отдельные специализированные части: формирование группы клеток зародыша — инициалей частей побега (квадрант) -»появление гипофизы в качестве инициальной клетки корня -> закладка примордиев семядолей с одновременным торможением делений клеток в участке между ними и появление прокамбия. Усложнение внутренних корреляционных взаимоотношений между клетками разных участков зародыша, по-видимому, указывает на становление собственной гормональной системы будущего организма.
Торпедовидная фаза (торпедо) развития зародыша (см. рис. 11.3) связана с делением клеток преимущественно поперек продольной оси и с более интенсивным ростом клеток в зачатках семядолей и в зоне гипокотиля. Отчетливее выделяются по вытянутой в длину форме клетки прокамбия в гипокотиле. Формируется промеристема корня. На этой фазе, кроме все более возрастающей концентрации ИУК и присутствия цитокинина, необходим гиббереллин (для роста гипокотиля). Изолированные зародыши в фазе торпедо удается выращивать на саха-розно-минеральной среде с добавлением витаминов и гиббереллина, в то время как для культивирования сердцевидных зародышей необходимо кокосовое молоко (эндосперм), в состав которого входят ауксин, цитокинины, неидентифицированные факторы углеводной природы и соединения, содержащие азот.
У некоторых двудольных удлиняющиеся семядоли и гипо-котиль изгибаются и складываются вдвое. Между семядолями закладывается апекс побега, происходит распад суспензора. Если семядоли выполняют функцию вместилища запасных веществ, то они заполняют почти весь объем созревающего семени, в них на последних этапах формирования зародыша откладываются запасные белки (в алейроновых зернах), крахмал (в амилопластах) и жиры (в сферосомах).
Приток питательных веществ в развивающиеся семязачатки (семяпочки) и затем в созревающие семена и формирующиеся плоды определяется тем, что эти участки становятся доминирующими центрами: в их тканях вырабатывается большое количество фитогормонов, прежде всего ауксина, в результате чего аттрагирующее действие этих тканей резко возрастает.
На последнем этапе созревания семена теряют значительное количество воды и у большинства видов средней полосы переходят в состояние покоя. Этот переход связан с уменьшением в тканях свободных (активных) ауксинов, цитокининов и гиббе-реллинов и с увеличением содержания АБК.
 Таким образом, в начале своего развития зародыши зависят от фитогормонов, поступающих из других тканей, прежде всего из эндосперма. Затем они сами начинают синтезировать фи-тогормоны, а при формировании семян ауксин выделяется из них в окружающие ткани, активируя рост околоплодника.
Таким образом, в начале своего развития зародыши зависят от фитогормонов, поступающих из других тканей, прежде всего из эндосперма. Затем они сами начинают синтезировать фи-тогормоны, а при формировании семян ауксин выделяется из них в окружающие ткани, активируя рост околоплодника.
Этап молодости у семенных растений начинается с прорастания семян или органов вегетативного размножения (например, клубней) и характеризуется быстрым накоплением вегетативной массы. Растения в этот период не способны к половому размножению.
В ювенильный период осуществляются прорастание семян (или вегетативных зачатков) и формирование вегетативных органов. Прорастание в свою очередь делится на фазы набухания семян, проклевывания, гетеротрофного роста проростка, перехода к автотрофному способу питания.
По окончании у семян периода покоя поглощение ими воды служит пусковым фактором прорастания. Это поглощение осуществляется благодаря повышению в ряде случаев проницаемости семенных покровов для воды и за счет гидратации биополимеров в клетках. В результате развивается онкотическое давление (давление набухания) и семенные покровы разрываются. Набухание практически не зависит от температуры, содержания О2, освещения.
 Проклевывание начинается, когда семена достигают критической влажности (40 — 65% в пересчете на сырую массу), и происходит путем роста растяжением самого зародышевого корня или гипокотиля, в результате чего кончик корешка выталкивается из семени. Деление клеток наступает обычно позже. Рост растяжением клеток зародышевой оси, возможно, обусловлен снижением содержания АБК при набухании семян. Выход корня обеспечивает закрепление прорастающего семени в почве и улучшает поглощение воды.
Проклевывание начинается, когда семена достигают критической влажности (40 — 65% в пересчете на сырую массу), и происходит путем роста растяжением самого зародышевого корня или гипокотиля, в результате чего кончик корешка выталкивается из семени. Деление клеток наступает обычно позже. Рост растяжением клеток зародышевой оси, возможно, обусловлен снижением содержания АБК при набухании семян. Выход корня обеспечивает закрепление прорастающего семени в почве и улучшает поглощение воды.
Вслед за корнем начинается рост побега. Прорастая в темноте (в почве), и корень, и побег ориентируются прежде всего по гравитационному вектору, при этом корень растет по направлению к центру Земли, т. е. углубляется в почву, а побег — от центра, т. е. направляется к свету. Рост осевых частей зародыша и проростка поддерживается фитогормонами. Причем у злаков ИУК и цитокинины вначале поступают в зародыш из эндосперма, ГА освобождаются из связанного состояния в зародышевой оси, а через несколько часов индуцируется синтез гиббереллинов в щитке. Эпителиальные клетки щитка начинают переваривать запасные вещества в эндосперме. К процессу пищеварения подключаются клетки алейронового слоя, активированные гиббереллином (см. 7.4). Подкисление тканей эндосперма за счет работы Н + -помп создает условия для кислого пищеварения и для всасывания продуктов разложения крахмала и белков клетками эпителия (симпорт Сахаров и аминокислот с ионами Н+ через плазмалемму). Клетки эпителия начинают вытягиваться и врастать в эндосперм, осуществляя
гетеротрофный способ питания растущего зародыша и проростка.
Рост зародышевого корня сопровождается появлением вдоль него зон деления, растяжения и дифференциации клеток (см. рис. 5.10). Корень сам начинает синтезировать цитокинины и ГА, которые направляются в побег. Побег удлиняется благодаря растяжению гипокотиля (у бобов, тыквы и др.) или мезо-котиля (у злаков). Листья не развиваются и гипокотиль в своей верхней части сильно изгибается наподобие крючка, что облегчает его перемещение в почве. В почечке у двудольных и в верхушке колеоптиля у злаков синтезируется ИУК.
Когда этиолированный побег (см. 11.5) достигает поверхности земли, возникают светоростовая и фотоморфогенетическая реакции: рост гипокотиля или мезокотиля резко подавляется, усиливается рост эпикотиля (первого настоящего междоузлия) и листьев. Содержание этилена в зоне крючка снижается и крючок распрямляется. Растение зеленеет и переходит к фо-тотрофному типу питания.
Благодаря дальнейшему росту главного, боковых и придаточных корней и формированию побегов за счет увеличения числа метамеров, ветвления, роста листовых пластинок, утолщения стебля растение к концу ювенильного этапа накапливает значительную вегетативную массу.
Продолжительность ювенильного периода у разных видов растений неодинакова: от нескольких недель (однолетние травы) до десятков лет (у древесных). Проростки по многим параметрам не похожи на взрослые растения. Различия часто хорошо заметны в форме листьев (хлопчатник, огурец, ипомея и др.). У некоторых растений наблюдаются различия и во внутреннем строении. Так, у проростков папоротника проводящая система организована проще, чем у взрослых растений. У юве-нильных растений менее мощная верхушечная меристема. Морфологическим признаком ювенильности в ряде случаев может служить также характер роста растения. Так, плющ в ювенильном состоянии представляет собой лазящий кустарник, а при переходе к взрослому состоянию образует вертикальный куст.
Этап молодости характеризуется полным отсутствием цветения или цветение слабо выражено даже при самых благоприятных для этого условиях. Здесь ярко проявляется роль компетенции, т. е. готовности специфически реагировать на то или иное индуцирующее воздействие. В данном случае юве-нильное растение не обладает компетенцией к факторам, вызывающим закладку органов полового или вегетативного размножения. Это может быть связано с отсутствием в органах-мишенях, воспринимающих гормоны, белков-рецепторов, участвующнх в индукции генеративного развития.
Однако отсутствие цветения само по себе не может служить показателем ювенильности, так как многие растения, находясь в зрелом состоянии, долго не зацветают без необходимых для нильности служат описанные выше морфологические признаки, например форма листьев.
 Для ювенильных растений характерна значительно большая способность к корнеобразованию, что с давних пор используется в практике садоводства. Предполагается, что лучшая укоре-няемость ювенильных черенков — следствие более высокого содержания в них ауксинов. Однако обработка ауксином взрослых черенков не вызывает возврата к ювенильному состоянию по морфологическим признакам, хотя и усиливает способность к корнеобразованию.
Для ювенильных растений характерна значительно большая способность к корнеобразованию, что с давних пор используется в практике садоводства. Предполагается, что лучшая укоре-няемость ювенильных черенков — следствие более высокого содержания в них ауксинов. Однако обработка ауксином взрослых черенков не вызывает возврата к ювенильному состоянию по морфологическим признакам, хотя и усиливает способность к корнеобразованию.
Ювенильное состояние поддерживается специфическим соотношением гормонов. В ряде случаев (например, у плюща) обработка гиббереллином приводит к образованию ювенильных листьев или к продлению ювенильного состояния.
Переход от ювенильного типа роста к зрелому наиболее хорошо заметен у древесных растений. У бука, например, возможно сосуществование всех фаз развития от ювенильности до зрелости. Это свойство древесных растений широко используется в садоводстве, когда для получения ювенильных растений отбирают черенки от основания сеянца, а черенки, взятые из верхних, более зрелых ярусов, укореняют или прививают для получения быстро зацветающих и плодоносящих растений.
Этап зрелости и размножения, т. е. период готовности к зацветанию и образованию органов вегетативного размножения, период закладки и роста органов размножения, формирования семян и плодов имеет особое значение в жизни растений и будет рассматриваться в гл. 12. Этап размножения растений в физиологическом плане изучен подробнее, чем другие этапы онтогенеза. В этот период растение наиболее жизнеспособно, оно сформировало вегетативную массу, достаточную для обеспечения роста и развития цветков, семян и плодов, которые являются гетеротрофными органами.
Этап старости и отмирания включает в себя период от полного прекращения плодоношения до естественной смерти организма. Это период прогрессирующего ослабления жизнедеятельности.
Продолжительность жизни растений, принадлежащих к разным таксономическим группам и различным жизненным формам, варьирует в очень широких пределах:
Секвойя.... 5000 лет Костер безостый 10-30 лет
Дуб...... 1500» Клевер.... 2-5»
Ель...... 1200» Двулетники (ка-
Липа..... 1000» пуста и др.) 2 года
Сосна..... 500» Однолетники
Виноград.... 80—100» (многие, травы) I год
Тау-сагыз... 100» Эфемеры... 2-4 недели
Это свидетельствует о генетической детерминированности нормы времени жизни каждого вида.
Старение и смерть — завершающие фазы онтогенеза любого многоклеточного растения, но термин «старение» может быть применим не только к целому растению, но и к отдельным его органам — листьям, плодам, частям цветков. Этот вид старения изучен наиболее полно, в то время как механизмы старения целого растения исследованы недостаточно.
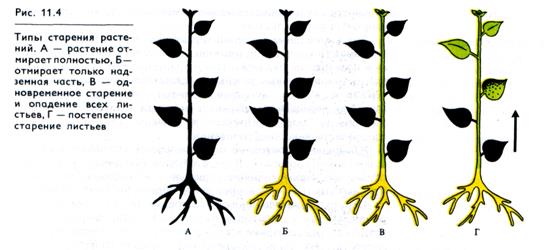
Для растений характерны разные типы старения (рис. 11.4). Так, однолетние растения отмирают целиком. У многолетних трав ежегодно полностью отмирает надземная часть, а корневая система и другие подземные части остаются жизнеспособными. У многих растений в процессе роста стареют и отмирают ранее образовавшиеся нижние листья. У листопадных деревьев осенью одновременно стареют и опадают все листья.
В процессе старения в листьях снижается содержание хлорофилла, белков, нуклеиновых кислот, падает интенсивность фотосинтеза, деградируют внутриклеточные органоиды. Одновременно активируются гидролитические ферменты, что на заключительном этапе старения приводит к автолизу.
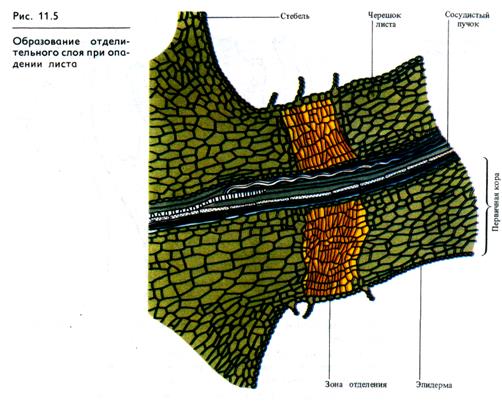
Процесс старения органа завершается его опадением. Перед опадением листа или плода в основании черешка или плодоножки образуется отделительный слой, представляющий собой зону, состоящую из клеток, ориентированных под прямыми углами к оси черешка (рис. 11.5). Размягчаются и частично растворяются клеточные стенки или срединные пластинки клеток отделительной зоны. Этот процесс индуцируется этиленом, вырабатываемым стареющими листьями.
Старение изолированных листьев задерживается при обработке их цитокинином, который стимулирует синтез РНК, белков и аттрагирующую активность клеток (см. рис. 2.8 и 2.9). У некоторых растений задержка старения может вызываться также ауксином и гиббереллинами. Абсцизовая кислота и этилен, наоборот, ускоряют процессы старения.
Одной из первых гипотез, объясняющих механизм старения целого растения, была гипотеза немецкого физиолога растений Г. Молиша, основанная на том, что у монокарпических растений — однолетних, двулетних (например, капуста) и некоторых
многолетних (агава, бамбук), цветущих лишь один раз, старение наблюдается сразу после цветения и созревания плодов. По мнению Молиша, старение вызывается оттоком большей части питательных веществ к развивающимся репродуктивным органам и отмирание наступает от истощения. Эта точка зрения подтверждается тем, что срезание цветков может задержать или даже предотвратить старение целого растения. Против этой гипотезы, однако, говорит тот факт, что удаление мужских цветков у двудомных растений также задерживает старение, хотя значительного оттока ассимилятов к цветкам в данном случае не происходит. Кроме того, у некоторых растений старение индуцируется определенным фотопериодом. Возможно, старение вызывается нарушением соотношения фитогормонов, возникающим как под действием внешних стимулов, так и внутренних факторов.
По мнению В. О. Казаряна (1959), основную роль в процессах старения играет функциональная корреляция между корнями и листьями: причиной ослабления роста и наступления старения может быть отставание развития корневой системы. Согласно этой концепции старение во время формирования плодов определяется угнетением роста корней из-за прекращения поступления в них ассимилятов. Уменьшение активности корней приводит к нарушению водного режима, фотосинтеза, синтеза белков и т. д., что снижает общую жизнедеятельность растений.
 У поликарпических растений, цветущих много раз, например у деревьев, по мере увеличения высоты ствола связь между листьями и корнями также ухудшается. Можно думать, что угнетение роста корней, чем бы это ни было вызвано, приводит к уменьшению поступления цитокинина в надземные органы и снижению пролиферативной активности апикальных меристем побега, что и служит причиной старения целого растения.
У поликарпических растений, цветущих много раз, например у деревьев, по мере увеличения высоты ствола связь между листьями и корнями также ухудшается. Можно думать, что угнетение роста корней, чем бы это ни было вызвано, приводит к уменьшению поступления цитокинина в надземные органы и снижению пролиферативной активности апикальных меристем побега, что и служит причиной старения целого растения.
Старение имеет большое биологическое значение. Оно является одним из способов адаптации растений к неблагоприятным условиям внешней среды. Кроме того, старение, по-видимому, способствует более быстрой эволюции, так как ускоряет смену поколений, т. е. «оборачиваемость» генетического материала.
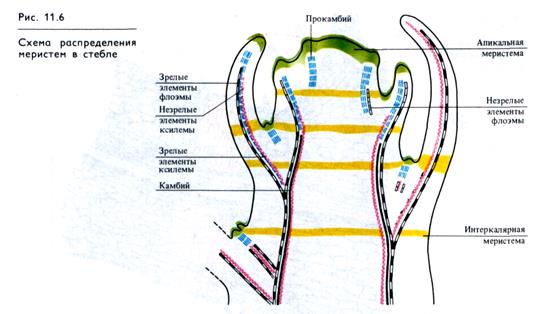
Растения в отличие от животных в течение всей жизни растут, образуя новые ткани и органы, которые закладываются в эмбриональных зонах — меристемах, где все клетки делятся. Существование меристем поддерживается инициальными клетками (инициалями), способными к делению неопределенно длительное время. Апикальные (верхушечные) меристемы расположены на концах побегов и корней. Латеральные (боковые) меристемы образуют слои клеток вдоль каждого побега и корня (у двудольных растений). К ним относятся первичные латеральные меристемы — прокамбий и перицикл и вторичные — камбий и феллоген. В основании молодых междоузлий и листьев функционируют интеркалярные (вставочные) меристемы (рис. 11.6). При повреждении у растений активируются меристемы, за счет которых регенерируют ткани, органы и целые организмы.
 После заложения тканей и органов в меристемах их рост происходит благодаря делению и последующему растяжению клеток. Форма и размеры органов и растения в целом определяются количеством и локализацией делящихся и растягивающихся клеток, интенсивностью и ориентацией их деления и роста. В процессе роста тканей и органов клетки, окруженные клеточными стенками и связанные плазмодесмами, не перемещаются относительно друг друга (согласованный рост). Эта особенность также отличает рост растительных организмов от животных, у которых рост зародыша сопровождается перемещением клеток и целых клеточных слоев.
После заложения тканей и органов в меристемах их рост происходит благодаря делению и последующему растяжению клеток. Форма и размеры органов и растения в целом определяются количеством и локализацией делящихся и растягивающихся клеток, интенсивностью и ориентацией их деления и роста. В процессе роста тканей и органов клетки, окруженные клеточными стенками и связанные плазмодесмами, не перемещаются относительно друг друга (согласованный рост). Эта особенность также отличает рост растительных организмов от животных, у которых рост зародыша сопровождается перемещением клеток и целых клеточных слоев.
Таким образом, морфогенез, т. е. формообразование у растений, включает в себя процессы заложения, роста и развития клеток (цитогенез), тканей (гистогенез) и органов (органогенез), которые генетически запрограммированы и скоординированы между собой. Апикальные меристемы побега и корня представляют собой не только образовательные ткани, из которых формируется весь растительный организм, но и главные координирующие (доминирующие) центры, влияющие на морфогенети-ческие процессы в целом растении (см. рис. 2.13).
Основные части побега — стебель, листья, почки, цветки и др. — закладываются в апикальной меристеме побега, которая является производной эмбриональной ткани дистального конца зародыша.
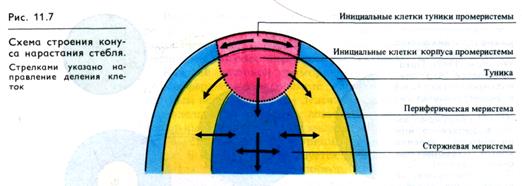
Апекс побега. Апекс (конус нарастания, точка роста) веге-тирующего побега семенного растения состоит из меристема-тических клеток, которые по своему размеру, частоте и направленности делений, по особенностям метаболизма могут быть разделены на несколько зон, прежде всего на тунику и корпус (рис. 11.7). Туника, или мантия,—это один, два или более слоев клеток, покрывающих апекс снаружи. Клетки туники делятся преимущественно антиклинально (т. е. плоскость деления перпендикулярна поверхности апекса). Из наружного слоя туники образуется эпидермис. Все остальные клетки, лежащие под туникой, входят в состав корпуса, в котором при более дробном анатомо-физиологическом разделении апикальной меристемы побега на зоны выделяют центральную, периферическую и стержневую меристемы (рис. 11.7). Дистальная группа клеток в тунике и центральная (аксиальная) зона выполняют функцию инициалей.
Клетки этих участков апекса относительно крупные и делятся сравнительно редко. Периферическая зона (инициальное кольцо) состоит из мелких меристематических клеток, которые интенсивно делятся. В них число рибосом выше, чем у инициальных клеток. Клетки этой зоны формируют при-мордии (зачатки) боковых органов побега — листьев и почек. Граница между туникой и корпусом в этой зоне исчезает. Клетки всех зон апекса имеют крупные ядра, густую цитоплазму и не содержат вакуолей.
Стержневая (колончатая) зона состоит из вакуолизиро-ванных клеток с относительно низким содержанием РНК. Клетки этой зоны делятся главным образом антиклинально и дают начало продольным рядам клеток первичной коры и сердцевины стебля. Границы между описанными зонами в апексе побега весьма условны и далеко не всегда различимы.
Конус нарастания побега, обладая высокой способностью к саморазвитию, тем не менее нуждается в притоке не только питательных веществ, но и фитогормонов. Изолированные апексы с двумя-тремя листовыми примордиями нормально развиваются лишь в том случае, если в питательной инкубационной среде присутствует цитокинин, а в ряде случаев — и ауксин.
Рост и развитие листа. Формирующийся лист проходит четыре фазы: 1) образование примордия, 2) формирование оси листа (черешка), 3) закладка пластинки листа за счет боковой меристемы, 4) рост пластинки растяжением.
Каждый листовой примордий образуется в виде бугорка в периферической меристеме апекса побега благодаря локальным периклинальным делениям клеток (плоскость деления параллельна поверхности апекса). У многих видов периклинальные деления в зоне закладки примордия происходят и в тунике. Примордий пазушной почки появляется несколько позже. В нем затем формируется апикальная меристема, гомологичная апексу главного побега.
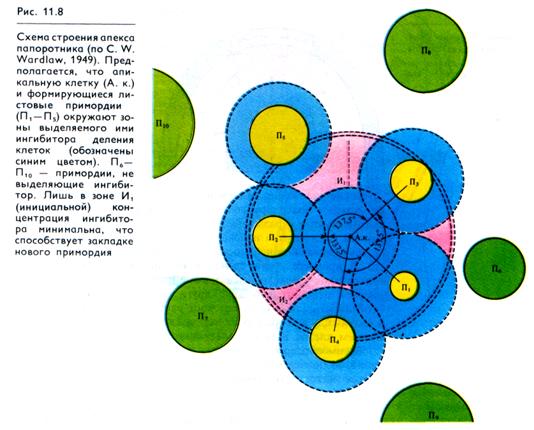
Период времени между заложением двух листовых зачатков называется пластохроном. Продолжительность его у разных видов и даже у одного и того же вида при разных условиях сильно варьирует: от нескольких часов до нескольких суток. Примордий листьев образуются на апексе в строго заданной последовательности, предопределяя расположение листьев на зрелом побеге, или филлотаксис. У растений распространен спиральный филлотаксис. Замечено, что на апексах с многочисленными примордиями угол между ними близок к значению 137,5°. При таком угле в идеальном случае ни один лист на стебле не располагается точно под другим, что обеспечивает их минимальную затененность. По теории В. Гофмейстера (1865), такое листорасположение достигается путем возникновения новых листовых зачатков в промежутках между уже существующими примордиями («теория доступного пространства»).
Согласно «теории отталкивания», предложенной Ю. Шоуте (1913), когда детерминируется центр листового примордия, в нем продуцируются специфические вещества, ингибирующие образование новых центров в непосредственной близости от заложившегося. Соответственно новый примордий развивается вне ингибиторных полей соседних.
Эти гипотезы довольно хорошо согласуются друг с другом, так как «доступное пространство» может быть обусловлено не только поверхностной зоной между примыкающими примордиями, но и зоной, свободной от их ингибирующего влияния. В этом месте и закладывается новая инициаль примордия (рис. 11.8). Действительно, если изолировать зону Ht двумя радиальными надрезами на апексе, то развитие здесь нового примордия ускоряется. Возникшие зачатки листьев оказывают влияние на нижележащие ткани, индуцируя дифференциацию проводящих пучков (рис. 11.6). Это действие обусловлено выделением ауксина, который синтезируется в формирующихся примордиях.
Верхушечные клетки конуса листового примордия делятся особенно интенсивно, превращая бугорок в пальцеобразный выступ (длина до 1 мм). Этот выступ состоит в основном из клеток будущей средней жилки и черешка листа. На краях зоны средней жилки начинает функционировать маргинальная (краевая) меристема, дающая начало листовой пластинке
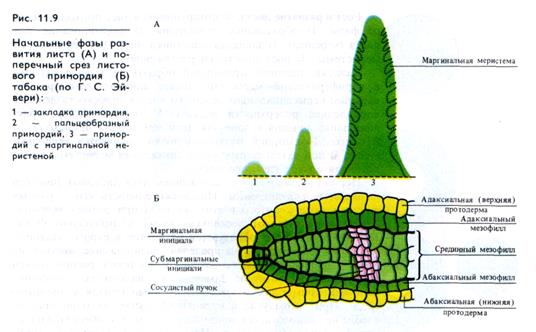

(рис. 11.9). Одновременно с этим прекращается верхушечный рост листа.
Инициальные клетки маргинальной меристемы и клетки самой этой меристемы делятся главным образом антиклинально, что приводит к увеличению площади листовой пластинки, а не ее толщины. Поверхностные инициальные маргинальные клетки образуют эпидермис, а субмаргинальные инициальные клетки — внутренние ткани листа (рис. 11.9).
После 8 — 9 циклов делений клетки маргинальной меристемы переходят к растяжению. Эпидермальные клетки кончают делиться первыми, но продолжают расти растяжением. Клетки губчатой паренхимы прекращают делиться и расти раньше других тканей. Поэтому продолжающееся деление и растяжение палисадной паренхимы и растяжение эпидермиса приводят к тому, что губчатые клетки отодвигаются друг от друга, образуя большие межклетники. Палисадные клетки делятся и растут со скоростью, близкой к росту эпидермиса. Этот процесс останавливается несколько раньше окончания растяжения эпидермиса. Поэтому и палисадные клетки несколько отрываются друг от друга, образуя небольшие межклетники.
 Особенность роста листа однодольных растений (например, злаков) состоит в том, что деления в возникшем на одной стороне апекса листовом бугорке распространяются в обе стороны и охватывают всю окружность стебля. Возникающий серповидный (у злаков) или кольцевой (у злаков) меристематический валик дает начало листу, растущему вверх. До формирования валика бугорок растет, как и у двудольных, верхушкой. Листовая пластинка удлиняется путем интеркалярного роста, который более продолжителен у основания пластинки.
Особенность роста листа однодольных растений (например, злаков) состоит в том, что деления в возникшем на одной стороне апекса листовом бугорке распространяются в обе стороны и охватывают всю окружность стебля. Возникающий серповидный (у злаков) или кольцевой (у злаков) меристематический валик дает начало листу, растущему вверх. До формирования валика бугорок растет, как и у двудольных, верхушкой. Листовая пластинка удлиняется путем интеркалярного роста, который более продолжителен у основания пластинки.
На рост листьев большое влияние оказывают периодичность, качество и интенсивность света. Свет сине-фиолетовой части спектра тормозит рост междоузлий и способствует росту листьев (у двудольных). Интенсивное освещение способствует развитию палисадной ткани.
Гормональная регуляция роста листа изучена недостаточно. Показано, что цитокинин и ауксин необходимы для формирования и развития примордиев и тканей листа, ауксин принимает участие в образовании жилок, гиббереллин способствует более интенсивному росту листовой пластинки в длину.
Рост листьев ограничен в отличие от пазушных почек, у которых заложенные апексы будут функционировать длительное время, если эти почки дадут боковые побеги.
Рост и развитие стебля. Стержневая меристема апекса и прокамбий, образование которого индуцируется растущими при-мордиями листьев, формируют основные ткани стебля. Выходя из меристематической зоны, клетки начинают растягиваться, что приводит к быстрому удлинению побега. Зона роста растяжением у побегов в отличие от корней достигает больших размеров (нескольких сантиметров). Рост растяжением стебля активируется гиббереллинами, стимулирующими переход большого количества клеток к этому типу роста, и ауксином, который непосредственно индуцирует удлинение клеток. Гиббе-реллины транспортируются из корня, но главным образом из листьев, и это позволяет регулировать скорость и продолжительность роста верхних междоузлий.
Стебель у двудольных утолщается за счет деятельности камбия, для активации которого необходима ИУК, поступающая из верхушки побега, а также благодаря пробковому камбию — феллогену, который образуется из различных слоев наружных клеток стебля.
Рост пазушных почек (ветвление) находится под двойным контролем: их рост и развитие ингибируют верхушечная почка побега и листья, в пазухах которых они находятся (см. 11.2.3).
Апекс корня. У высших растений апикальная меристема корня имеет сравнительно простое строение (см. рис. 5.10). Это зона длиной 1—2 мм. В ней не образуются боковые органы, как в верхушечной меристеме побега. Корневая меристема формирует ткани корня и корневой чехлик, защищающий корень при его продвижении в почве.
Кроме активно делящихся клеток, в меристеме корня находится группа клеток, расположенных между корневым чехли-ком и активной меристематической зоной, для которых характерны низкий уровень синтеза ДНК и очень редкие деления клеток. Например, длительность митотического цикла в клетках этой зоны у кукурузы 174-239 ч против 14 — 39 ч у окружающих меристематических клеток. Эта группа клеток получила название «покоящегося центра». Предполагается, что «покоящийся центр» является промеристемой активной апикальной меристемы корней, восстанавливающей численность быстро делящихся специализированных инициальных клеток при их естественном изнашивании или повреждении. В этом смысле функции клеток «покоящегося центра» сходны с аналогичной ролью центральной зоны («зоны ожидания») апекса побега.
 Одна группа инициальных клеток локализована на дисталь-ном конце апекса и продуцирует клетки ризодермы и корневого чехлика. Другая инициаль связана с репродуцированием клеток первичной коры. Третья ответственна за поддержание меристематической активности клеток, дифференцирующихся затем в различные клетки и ткани проводящего пучка. Образование рядов специализированных клеток в корне можно проследить непосредственно от их клеток-инициалей. Таким образом, апикальная меристема корня, как и апекс побега, продолжают ту ткане- и органообразующую деятельность, которая началась еще при формировании зародыша.
Одна группа инициальных клеток локализована на дисталь-ном конце апекса и продуцирует клетки ризодермы и корневого чехлика. Другая инициаль связана с репродуцированием клеток первичной коры. Третья ответственна за поддержание меристематической активности клеток, дифференцирующихся затем в различные клетки и ткани проводящего пучка. Образование рядов специализированных клеток в корне можно проследить непосредственно от их клеток-инициалей. Таким образом, апикальная меристема корня, как и апекс побега, продолжают ту ткане- и органообразующую деятельность, которая началась еще при формировании зародыша.
 Изолированные «покоящиеся центры» при культивировании на питательной среде нуждаются в присутствии ИУК и цито-кинина (0,1 мг/л). Изолированные кончики корней однодольных растут при добавлении ауксина, а у многих двудольных кончики корней развиваются и без экзогенных фитогормонов (среда: сахароза, минеральные вещества, витамины). Очевидно, в последнем случае ИУК синтезируется ба-зальной частью отрезка корня.
Изолированные «покоящиеся центры» при культивировании на питательной среде нуждаются в присутствии ИУК и цито-кинина (0,1 мг/л). Изолированные кончики корней однодольных растут при добавлении ауксина, а у многих двудольных кончики корней развиваются и без экзогенных фитогормонов (среда: сахароза, минеральные вещества, витамины). Очевидно, в последнем случае ИУК синтезируется ба-зальной частью отрезка корня.
Ростовые корреляции — это зависимость роста и развития одних органов или частей от других.
Самый простой тип корреляций связан с питанием. Гетеротрофные части, например корни, зависят в своем развитии от побега, снабжающего их органическими соединениями. В свою очередь развитие побега находится в зависимости от корня, поставляющего минеральные вещества и воду. Корреляции, связанные с питанием, наблюдаются в случае торможения вегетативного роста при плодоношении, что объясняется оттоком питательных веществ в развивающиеся семена и плоды.
Однако даже такие простые корреляции роста далеко не полностью удается объяснить с помощью теории конкуренции за доступное питание. Во многих случаях основную роль играют не столько трофические, сколько гормональные взаимодействия между частями растения. Одним из типичных примеров гормональной регуляции может служить апикальное доминирование — коррелятивное торможение верхушкой побега или корня развития соответственно пазушных почек или боковых корней. Этот пример корреляций изучен более полно, поэтому рассмотрим его подробнее.
Апикальная почка побега обычно растет значительно энергичнее, чем нижележащие пазушные почки, несмотря на то, что пазушные находятся, казалось бы, в лучшем положении по отношению к органическому и минеральному питанию, поступающему из зрелых листьев и корневой системы. Степень апикального доминирования может меняться с возрастом растения и в зависимости от вида. Как правило, апикальное доминирование снижается по мере старения и может поисчезать, в частности, в том случае, когда апикальная меристема образует цветок.
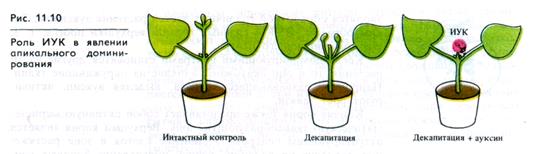
лностью К. Тиманн и Ф. Скуг (1934) показали, что экзогенная ИУК заменяет апикальную почку в ингибировании пазушных почек у растений бобов, что позже подтвердилось для большинства растений (рис. 11.10). По гипотезе «прямого ингибировани.ч» предполагалось, что верхушка побега снабжает боковые почки избыточным количеством ауксина, подавляя тем самым их рост и развитие. Однако определение содержания ауксина в боковых почках не подтверждает это предположение.
 Одной из распространенных точек зрения является трофическая теория, согласно которой апикальная почка так или иначе предотвращает доступ питательных веществ к боковым почкам, тормозя этим их развитие. Эта точка зрения подтверждается тем, что ИУК, нанесенная на декапитированную верхушку, задерживает образование проводящих элементов, соединяющих боковые почки с центральной проводящей системой; сама же декапитация стимулирует этот процесс. Тем не менее некоторые данные показывают, что пробуждение и начальный этап роста пазушных почек не зависят от существования проводящих пучков между ними и стеблем, хотя дальнейший рост почек может продолжаться только при условии связи между проводящими системами.
Одной из распространенных точек зрения является трофическая теория, согласно которой апикальная почка так или иначе предотвращает доступ питательных веществ к боковым почкам, тормозя этим их развитие. Эта точка зрения подтверждается тем, что ИУК, нанесенная на декапитированную верхушку, задерживает образование проводящих элементов, соединяющих боковые почки с центральной проводящей системой; сама же декапитация стимулирует этот процесс. Тем не менее некоторые данные показывают, что пробуждение и начальный этап роста пазушных почек не зависят от существования проводящих пучков между ними и стеблем, хотя дальнейший рост почек может продолжаться только при условии связи между проводящими системами.
В последнее время обнаружено, что нанесение кинетика на покоящуюся почку инициирует начальные этапы ее развития, причем дальнейший рост почки возможен только при постоянном снабжении ее кинетином, а некоторое время — и ауксином. Таким образом, роль ауксина в апикальной почке заключается в создании мощного аттрагирующего центра, в результате чего не только питательные вещества, но и цигокинин, синтезированный в корнях, поступают преимущественно в апикальную почку. Приток цитокинина к пазушным почкам после устранения апикальной усиливает в них клеточные деления. Формирующиеся в почках листовые зачатки начинают синтезировать ауксин, необходимый для дальнейшего стабильного развития боковых побегов.
 Развивающаяся верхушечная почка влияет также на рост клеток в зонах растяжения побега и корня, индуцирует формирование проводящих пучков. Ориентация листьев, боковых побегов и корней также зависит от функциональной активности верхушки (см. рис. 2.13). Все эти эффекты верхушки воспроизводятся обработкой различных частей растения ауксином. Таким образом, ИУК — главный гормон верхушки побега, т. е. его доминирующего центра.
Развивающаяся верхушечная почка влияет также на рост клеток в зонах растяжения побега и корня, индуцирует формирование проводящих пучков. Ориентация листьев, боковых побегов и корней также зависит от функциональной активности верхушки (см. рис. 2.13). Все эти эффекты верхушки воспроизводятся обработкой различных частей растения ауксином. Таким образом, ИУК — главный гормон верхушки побега, т. е. его доминирующего центра.
Когда доминирующими центрами становятся другие части растения, то и они оказывают влияние на окружающие ткани. Например, развивающиеся семена, выделяя ауксин, активируют рост завязи.
 Кончик корня также представляет собой активную меристе-матическую тканеобразующую зону. Верхушка корня является аттрагирующим центром. Удлинение клеток в зоне растяжения, закладка проводящих пучков, образование боковых корней тоже контролируются кончиком корня. Более того, от деятельности верхушки корня зависят индукция развития стеблевых апексов, рост и функциональная активность листьев (см. рис. 2.13).
Кончик корня также представляет собой активную меристе-матическую тканеобразующую зону. Верхушка корня является аттрагирующим центром. Удлинение клеток в зоне растяжения, закладка проводящих пучков, образование боковых корней тоже контролируются кончиком корня. Более того, от деятельности верхушки корня зависят индукция развития стеблевых апексов, рост и функциональная активность листьев (см. рис. 2.13).
Некоторые из этих явлений, находящихся под контролем верхушки корня, можно воспроизвести с помощью обработки цитокинином. Наряду с ИУК цитокинин необходим для формирования проводящих пучков, у двудольных он тормозит удлинение стебля. Цитокинин контролирует образование примор-диев листьев, их рост и трофику. Этот гормон синтезируется в кончике корня и с ксилемным током перемещается в верхушки побегов и в листья. Помимо цитокинина в верхушке корня (в корневом чехлике) образуется АБК, которая, возможно, наряду с ИУК участвует в регуляции роста корня.
Гормональное взаимодействие доминирующих центров побега (ИУК) и корня (цитокинин) служит, по-видимому, важнейшим эндогенным механизмом роста и морфогенеза в целом растении. К этому добавляется взаимодействие доминирующих центров с листьями, вырабатывающими гиббереллины и АБК. Возникновение градиентов количества и соотношения фитогормонов наряду с появлением или исчезновением компетентности различных тканей к конкретным фитогормонам представляют основу для пространственной организации роста и морфогенеза.
Ростовые корреляции широко используются в практике растениеводства для получения большего количества хозяйственно полезной продукции. Например, пасынкование — удаление боковых побегов у томатов — способствует образованию более крупных плодов; пикировка — обрывание концов корней при пересадке рассады овощей — создает условия для увеличения массы боковых и придаточных корней; вершкование — удаление соцветий (у табака) — для повышения выхода листьев и др.
Процессам роста, как и другим физиологическим явлениям, свойственна периодичность, которая обусловливается как особенностями самих процессов роста, так и факторами внешней среды.
У растений наиболее распространены так называемые циркадные ритмы с периодом около суток. С такой периодичностью изменяется, например, митотическая активность в меристемах (период около 20 ч) и скорость роста (период 24 ч).
 К подобным эндогенным ритмам относятся также ритмы фотосинтеза и дыхания, транспорта веществ, транспирации, открывания и закрывания цветков и т. д. Околосуточные ритмы тесно связаны с суточными колебаниями освещенности, температуры и других факторов среды, причем сложившаяся периодичность физиологических процессов некоторое время сохраняется у растений и при изменении условий среды, вследствие чего эти ритмы названы эндогенными. Благодаря эндогенным ритмам живые организмы хорошо приспособлены к тем условиям, в которых они обитают, мало завися от случайных погодных флуктуации.
К подобным эндогенным ритмам относятся также ритмы фотосинтеза и дыхания, транспорта веществ, транспирации, открывания и закрывания цветков и т. д. Околосуточные ритмы тесно связаны с суточными колебаниями освещенности, температуры и других факторов среды, причем сложившаяся периодичность физиологических процессов некоторое время сохраняется у растений и при изменении условий среды, вследствие чего эти ритмы названы эндогенными. Благодаря эндогенным ритмам живые организмы хорошо приспособлены к тем условиям, в которых они обитают, мало завися от случайных погодных флуктуации.
Кроме суточной периодичности рост растений подвержен изменениям в течение сравнительно длительных периодов, например сезонной периодичности. Такая периодичность выражается в образовании годичных колец в древесине растений умеренного пояса, у которых ростстволов в толщину, достигая максимума в летнее время, прекращается осенью.
Для роста растений на любых этапах онтогенеза характерен период покоя. Существует покой на этапе эмбрионального развития растений (покой зиготы и семян) и покой побегов растений, находящихся на разных фазах вегетативного развития. Семена могут оставаться в состоянии покоя длительное время, сохраняя жизнеспособность. Различают вынужденный покой, причиной которого являются факторы внешней среды, препятствующие прорастанию, и физиологический покой, под которым понимается задержка прорастания, вызванная свойствами зародыша или окружающих его тканей (эндосперма и семенной кожуры, а также околоплодника).
Причины физиологического покоя семян разнообразны и могут вызываться несколькими механизмами. Эндогенный покой определяется главным образом свойствами зародыша, т. е. его морфологической недоразвитостью, особым физиологическим состоянием или же сочетанием обеих этих причин. Состояние покоя регулируется балансом фитогормо-нов, стимулирующих и ингибирующих рост, — ИУК, цитокини-нов, гиббереллинов и АБК. В ряде случаев торможение роста зародышей связано со сверхоптимальным содержанием АБК и ИУК, а выход из состояния покоя — со снижением концентраций этих гормонов и с одновременным увеличением активности гиббереллинов и цитокининов.
Покой почек и побегов более, чем покой семян, определяется климатическими условиями, являясь приспособлением для перенесения неблагоприятных условий. У древесных растений к зиме апексы побега прекращают активный рост и покрываются почечной чешуей, превращаясь в зимние покоящиеся почки. В таком состоянии покоя растения гораздо морозоустойчивее, чем в состоянии активного роста. Однако причина, по которой покоящиеся почки более устойчивы, полностью не ясна. Возможно, это в какой-то мере связано с уменьшением потери воды. У многих растений этой же цели служат специальные покоящиеся органы — луковицы, клубни, клубнелуковицы и корневища. Эти органы могут быть адаптированы также к перегреву, летней засухе, как, например, луковицы многих пустынных растений.
 Покой побегов и почек так же, как и покой семян, можно подразделить на вынужденный (обусловленный неблагоприятными температурными и световыми условиями) и физиологический. Физиологический покой существует и у таких органов, как корневища, клубнелуковицы и клубни.
Покой побегов и почек так же, как и покой семян, можно подразделить на вынужденный (обусловленный неблагоприятными температурными и световыми условиями) и физиологический. Физиологический покой существует и у таких органов, как корневища, клубнелуковицы и клубни.
Весь период покоя почки в зависимости от его причин можно подразделить на три фазы. Летний покой (предпокой), т. е. отсутствие роста почки летом, обусловлен коррелятивным ингибированием. Затем эти почки у многих видов переходят в состояние глубокого зимнего покоя. В третий период постпокоя почки уже не находятся в состоянии внутреннего покоя, но в течение некоторого времени не могут расти из-за неблагоприятных температурных условий.
У большинства видов средней полосы состояние покоя контролируется фотопериодической реакцией: длинный день ускоряет вегетативный рост, а короткий приводит к прекращению роста и формированию покоящихся почек. Однако ряд плодовых деревьев и некоторые другие виды к изменениям длины дня сравнительно нечувствительны. Выход из покоя тем не менее не контролируется исключительно изменением длины дня. Для многих растений выход почек из покоя возможен только после длительного воздействия низкими температурами или в результате кратковременной обработки высокими температурами (тепловые ванны, 30-40°С).
При сравнении покоя почек и покоя семян обнаруживается сходство, особенно заметное в тех случаях, когда почки и семена относятся к одному и тому же виду. Это наводит на мысль, что природа покоя в обоих случаях одна и та же. Наибольшее сходство наблюдается при рассмотрении гормонального контроля покоя.
Покой почек и других покоящихся органов может быть преодолен с помощью тех же типов гормонов (гиббереллинов, ци-токининов и этилена), которые активны в прерывании покоя семян. Сходным образом у почек и семян снижается уровень эндогенных гиббереллинов и цитокининов при вхождении в состояние покоя. В том и другом случае показано, что веществом, индуцирующим покой, может быть АБК. Однако несмотря на множество экспериментальных данных, механизм действия фитогормонов в явлениях покоя почек и семян окончательно не выяснен.
 Регенерация — это восстановление организмом поврежденной или утраченной части тела. Способность к регенерации широко распространена в растительном мире от низших до высших таксонов, причем разнообразие форм регенерации очень велико. Это обусловлено следующими причинами. Во-первых, регенерация — один из основных неспецифических способов защиты растений, ведущих неподвижный, прикрепленный образ жизни, от всякого рода повреждений и травм. Во-вторых, многие формы регенерации успешно используются растениями как способ вегетативного размножения.
Регенерация — это восстановление организмом поврежденной или утраченной части тела. Способность к регенерации широко распространена в растительном мире от низших до высших таксонов, причем разнообразие форм регенерации очень велико. Это обусловлено следующими причинами. Во-первых, регенерация — один из основных неспецифических способов защиты растений, ведущих неподвижный, прикрепленный образ жизни, от всякого рода повреждений и травм. Во-вторых, многие формы регенерации успешно используются растениями как способ вегетативного размножения.
Явление регенерации представляет большой теоретический и практический интерес. Теоретический потому, что различные формы регенерации служат прекрасными модельными системами для изучения механизмов морфогенеза. Ведь лат. regeneratio — это возрождение, вторичное развитие. Причем у растений можно отдельно изучать восстановление тех или иных тканей, органов или даже целого организма. В практической деятельности человека регенерация используется издавна. Прежде всего это размножение черенками, которые у многих растений легко укореняются. Большой вклад в разработку проблемы регенерации у растений внесли советские исследователи Н. П. Кренке (1950), Р. X. Турецкая (1961), А. Г. Юсуфов (1981, 1982) и др.
В настоящее время не существует общепринятой классификации форм регенерации у растений, а также устоявшейся терминологии. Если исходить из механизмов регенерации, то классификацию наиболее часто встречающихся способов регенерации у растений можно представить следующим образом:
 I. Физиологическая регенерация.
I. Физиологическая регенерация.
II. Травматическая регенерация:
1. Регенерация, обусловленная дедифференцировкой клеток:
а) заживление ран;
б) органогенез, связанный с образованием каллуса;
в) соматический эмбриогенез;
г) восстановление частей без образования каллуса;
2. Регенерация на уровне меристем:
а) восстановление апикальных меристем;
б) органогенез из предсуществующих зачатков;
в) органогенез из новообразованных адвентивных зачатков.
Кратко рассмотрим каждый из перечисленных способов.
Физиологическая регенерация. При этом способе восстанавливаются части при их естественном изнашивании. Примеры физиологической регенерации — постоянное восполнение слущивающихся клеток корневого чехлика, замена старых элементов ксилемы новыми (у древесных), замена корки у стволов деревьев и др. В животных организмах аналогичным образом идет постоянное обновление клеток слизистой желудка, клеток кожи и т. д.
Заживление ран. Ткани, оказавшиеся на поверхности раны, дедифференцируются, их клетки начинают периклинально делиться и образуют феллоген, превращающийся в пробку. Поверхность раны может затягиваться также каллусной тканью.
Органогенез, обусловленный образованием каллуса. Начальный этап дедифференциации клеток на поверхности раны аналогичен тому, что происходит при заживлении ран. Однако клетки, дедифференцируясь, переходят к неорганизованному делению и возникает каллусная ткань, состоящая из рыхло соединенных друг с другом паренхиматиче-ских клеток. При определенных условиях (см. ниже) отдельные клетки или группы клеток могут дать начало адвентивным  (т. е. возникшим не из эмбриональных тканей) органам: корням, побегам, листьям. В естественных условиях каллус на поверхности среза стебля или корня обычно образуется из камбиальных клеток.
(т. е. возникшим не из эмбриональных тканей) органам: корням, побегам, листьям. В естественных условиях каллус на поверхности среза стебля или корня обычно образуется из камбиальных клеток.
Соматический эмбриогенез. Каллус на раневой поверхности образуется так, как описано выше. Из отдельных клеток каллуса, начинающих организованно делиться, формируются соматические зародыши (эмбриоиды), из которых при определенных условиях развивается целый организм. Такой процесс идет и в районе перерезанных жилок листа бегонии, где из единичных эпидермальных клеток образуются целые растеньица.
Восстановление частей без образования каллуса. Примером такого способа регенерации служит формирование адвентивных побегов из единичных эпидермальных клеток на некотором удалении от раневой поверхности. Другой пример — превращение паренхимных клеток коры в клетки ксилемы при образовании обходного участка проводящего пучка вокруг места его прерывания. Направление регенерации проводящих элементов определяется прежде всего полярным базипетальным транспортом ауксина, который индуцирует генетическую программу ксилемообразования. Дифференцировке элементов флоэмы наряду с присутствием ИУК и цитокинина способствует высокая (4-8%) концентрация сахарозы.
Другие пути восстановления утраченных частей у растений связаны с деятельностью апикальных или латеральных меристем.
Восстановление апикальных меристем. При продольном рассечении конуса нарастания из каждой половины могут регенерировать отдельные апексы. Конус нарастания как побега, так и корня регенерирует при удалении небольшого участка его дистального конца (не более 80 мкм). У развивающегося молодого листа папоротника восстанавливается отрезанная меристематическая верхушка.
Органогенез из предшествующих зачатков. Восстановление надземных органов у высших растений осуществляется, как правило, за счет отрастания покоящихся (пазушных) почек при устранении доминирующего влияния апикальной почки побега (см. 11.2.3). Повреждение или частичное удаление дистальной части корневой системы также способствует росту зачатков боковых корней вследствие устранения тормозящего действия кончика корня. Такой способ восстановления утраченных частей присущ только растениям.
Органогенез из новообразованных адвентивных зачатков. Стеблевые черенки многих древесных травянистых растений образуют корни благодаря активации периклинальвых делений в камбии или перицикле, выполняющих функции латентных меристем. Индукция делений клеток связана с действием ИУК, которая, перемещаясь базипетально, накапливается в нижней части черенка.
Таким образом, растения могут восстанавливать как надземные, так и подземные части даже при полной их утрате. Для этого используются те или иные вышеописанные способы регенерации или их комбинации.
Выяснение принципов, лежащих в основе образования специфической формы каждого организма и его частей,—наиболее трудная и еще мало изученная проблема биологии. Для ее решения применительно к многоклеточным организмам необходимо прежде всего понять, каким образом клетки, ткани и органы взаимодействуют между собой в ходе онтогенеза. В гл. 2 отмечалось, что межклеточные системы регуляции у растений включают в себя гормональные, электрические и трофические факторы, которые оказывают влияние на генетическую, мембранную и метаболическую регуляторные системы в каждой клетке. Для процессов эмбриогенеза и регенерации, в основе которых лежат одни и те же принципы, наиболее важны последовательная индукция определенных генетических программ и морфофизиологическая ориентация в пространстве. Запуск генетических программ осуществляется специфическими химическими и физико-химическими факторами, а ориентация в пространстве создается полярностью, в основе которой лежат прежде всего мембранные процессы. Клетки постоянно получают сигналы из внешней и внутренней среды (тестируют свое положение и состояние). В соответствии с этим корректируются их функциональная активность, считка генетической информации и векторы поляризации. Такая корректировка получила название «эффекта положения».
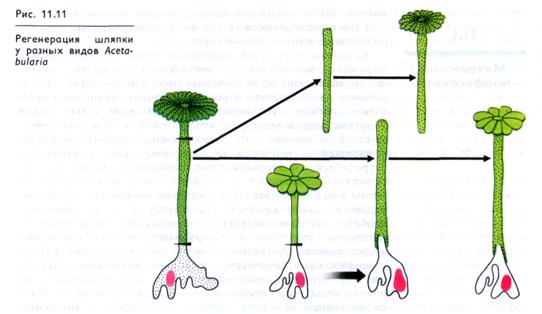
 Среди растений есть прекрасные объекты для изучения генетических аспектов формообразования. У морской одноклеточной водоросли ацетабулярии (порядок сифоновые), достигающей в длину 5 — 6 см, ядро находится в одном из ризоидов, что позволяет легко его удалять. Клетка ацетабулярии состоит из трех частей: ризоидного основания, стебелька и спорообразую-щей шляпки, по строению которой различаются виды. При отрезании шляпки оставшаяся часть клетки легко ее регенерирует. Шляпка регенерирует даже в том случае, когда отрезается ризоид, содержащий ядро.
Среди растений есть прекрасные объекты для изучения генетических аспектов формообразования. У морской одноклеточной водоросли ацетабулярии (порядок сифоновые), достигающей в длину 5 — 6 см, ядро находится в одном из ризоидов, что позволяет легко его удалять. Клетка ацетабулярии состоит из трех частей: ризоидного основания, стебелька и спорообразую-щей шляпки, по строению которой различаются виды. При отрезании шляпки оставшаяся часть клетки легко ее регенерирует. Шляпка регенерирует даже в том случае, когда отрезается ризоид, содержащий ядро.
Форма регенерирующей шляпки определяется ядром: при пересадке стеблевой части Acetabularia mediterranea на основание клетки A. wettsteinii, содержащее ядро, регенерирует шляпка формы, характерной для A. wettsteinii. Отрезок же стеблевой части без ядра регенерирует «свою» шляпку (рис. 11.11).
Эти и другие опыты показывают, что в клеточном ядре образуются специфические для каждого вида долгоживущие морфогенетические информационные РНК, поступающие в стебелек и «управляющие» морфогенезом шляпки через синтез специфических белков. Это доказывают и опыты с иснользова-нием ингибиторов синтеза белков и нуклеиновых кислот, предотвращающих регенерацию шляпки.
У многоклеточных растений механизмы внутриклеточной генной регуляции не отличаются от аналогичных процессов в клетках ацетабулярии, однако включение или выключение генетических программ в каждой клетке зависит от поступления сигналов из других клеток, тканей и органов. При изучении фи-зиолого-биохимических основ морфогенеза у высших растений большое распространение получила культура тканей (экспериментальный метод выращивания тканей вне организма), являющаяся более простой системой, чем культура изолированных органов или интактное, т. е. неповрежденное, растение. Возможность изучения формообразовательных процессов в культуре тканей определяется тем, что каждая растительная клетка тотипотентна. Этот метод дает прекрасную возможность изучать действие трофических и гормональных факторов на процессы морфогенеза.

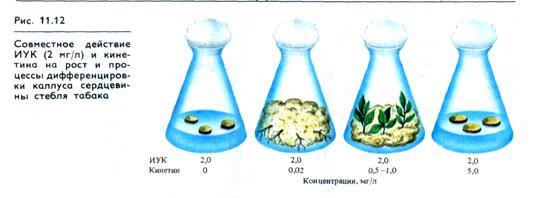
Например, одна ИУК (0,5 — 2 мг/л) вызывает у паренхимных клеток сердцевины клеток табака лишь рост растяжением. Ф. Скуг (1957) показал, что при добавлении в питательную среду вместе с ауксином кинетика (0,02—1 мг/л) индуцируется деление клеток и образуется рыхлая каллусная ткань (рис. 11.12). Если концентрация ИУК сравнительно высока (2 мг/л), а кинетина низка (0,02 мг/л), то в каллусной ткани индуцируется образование корней. Если же концентрация кинетина повышается до 0,5—1 мг/л, то корне-образование подавляется, а в каллусной ткани наблюдается закладка и рост стеблевых почек. В присутствии высоких концентраций ауксина и кинетина (5 мг/л) рост каллуса и процессы органообразования подавлены. Один кинетин на ткани сердцевины стебля табака видимого влияния не оказывает. Таким образом, опыты показывают, что преобладание ауксина необходимо для включения генетической программы корнеобразо-вания, а высокое содержание цитокинина в присутствии ИУК индуцирует функциональную активность генов, ответственных за программу побегообразования.
 Иные специфические сочетания фитогормонов, трофических факторов и физико-химических условий индуцируют другие генетические программы. Такое морфогенетическое воздействие осуществляется лишь в том случае, если клетки компетентны (чувствительны) к поступающим сигналам (эффекторам). Гипотетический механизм избирательного включения генов с участием эффекторов представлен на рис. 2.3.
Иные специфические сочетания фитогормонов, трофических факторов и физико-химических условий индуцируют другие генетические программы. Такое морфогенетическое воздействие осуществляется лишь в том случае, если клетки компетентны (чувствительны) к поступающим сигналам (эффекторам). Гипотетический механизм избирательного включения генов с участием эффекторов представлен на рис. 2.3.
Важнейшее условие формообразования при развитии организма — поляризация биологических структур. Под полярностью подразумевают специфическую ориентацию процессов и структур в пространстве, приводящую к появлению морфо- физиологических градиентов. Полярность определяет положений осей, обусловливающих форму клеток, органов и целого организма.
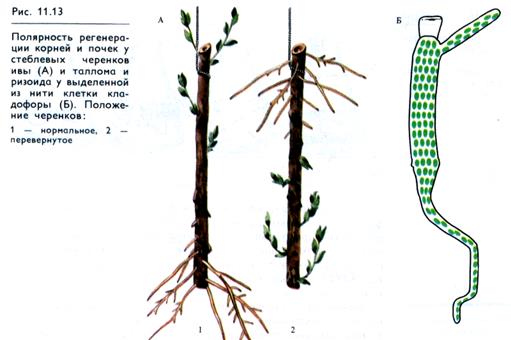
Полярность особенно наглядно представлена у растений, для которых характерна биполярная структура (главная ось: побег — корень). В физиологическом плане полярность проявляется у растений, в частности в процессах регенерации. У стеблевых и корневых черенков независимо от их положения в пространстве побеги развиваются с морфологически апикального (по отношению к верхушке стебля), а корни — с базального концов (рис. 11.13, А). Это объясняется тем, что ИУК, перемещаясь полярно, скапливается в морфологически нижнем конце черенка (см. 11.3) и индуцирует включение генетической программы корнеобразования.
Свойство полярности присуще не только организму в целом и его органам, но и отдельным клеткам. Например, у изолированных клеток нитчатой зеленой водоросли Cladophora ризоид образуется из морфологически нижнего конца клетки, а таллом — из верхнего конца (рис. 11.13, Б). С помощью цитофотометрии биохимическая полярность клеток показана у протонемы мха (О. Т. Демкив, 1974).
Однако полярность не является изначальным и неизменно существующим свойством биологических объектов. У спор хвощей и папоротников полярность возникает лишь после определенных внешних воздействий, например, в условиях односторонне падающего света. При делении такой поляризованной споры освещенная сторона и соответствующая дочерняя клетка формируют заросток, а затененная — ризоид.
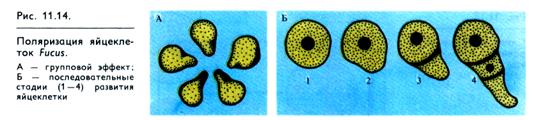
Механизм поляризации особенно подробно изучен у яйцеклетки бурой морской водоросли Fucus. До оплодотворения яйцеклетка фукуса лишена оболочки, ядро расположено в центре клетки и вначале не наблюдается сколько-нибудь заметной полярности в ее строении. После оплодотворения клетка опускается на дно, покрывается оболочкой и через некоторое время на ее нижней поверхности начинается образование ризоид-ного выступа. Первое деление яйцеклетки проходит в направлении, перпендикулярном образовавшейся оси. Верхняя клетка дает начало большей части таллома, нижняя — небольшой части таллома и ризоиду (рис. 11.14, Б). По-видимому, сила гравитации в данном случае не представляет собой определяющего фактора в индуцировании полярности, так как при развитии яйцеклеток фукуса в темноте ризоиды могут расти в различных направлениях. При одностороннем освещении ризоид образуется с затененной стороны. Наложение слабого электрического поля также вызывает поляризацию яйцеклетки, причем выступ ризоида появляется со стороны анода. Локальная обработка ауксином или односторонний вход в яйцеклетку Са2+ индуцирует образование ризоида в месте обработки. Все эти и другие способы воздействия приводят к возникновению локальных электрических токов (поляризации). Кальций индуцирует секреторную активность яйцеклетки и локальное выделение полисахаридов клеточной стенки, что и способствует появлению ризоидного выступа. У ядра образуются пальцео бразные выросты ядерной оболочки, направленные в сторону будущего ризоида, и в этом же участке цитоплазмы концентрируются митохондрии и рибосомы.
 Предполагается, что вследствие электрической поляризации яйцеклетки в ее плазмалемме происходит латеральное электро- форетическое перемещение липопротеиновых компонентов с положительным или отрицательным зарядом (L. F. Jaffe et al. 1977—1980). Эти компоненты (ионные каналы, насосы, ферменты и др.) затем закрепляются на полюсах клетки с помощью микрофиламентов и микротрубочек цитоскелета, что необратимо фиксирует возникшую первичную поляризацию и определяет главную ось тела растения. При последующем делении яйцеклетки (плоскость деления перпендикулярна оси поляризации) ядра в дочерних клетках попадают в совершенно разные условия, возникшие в поляризованной цитоплазме, и вследствие этого начинают поставлять неидентичную генетическую информацию. Таким образом происходит дифференциация клеток.
Предполагается, что вследствие электрической поляризации яйцеклетки в ее плазмалемме происходит латеральное электро- форетическое перемещение липопротеиновых компонентов с положительным или отрицательным зарядом (L. F. Jaffe et al. 1977—1980). Эти компоненты (ионные каналы, насосы, ферменты и др.) затем закрепляются на полюсах клетки с помощью микрофиламентов и микротрубочек цитоскелета, что необратимо фиксирует возникшую первичную поляризацию и определяет главную ось тела растения. При последующем делении яйцеклетки (плоскость деления перпендикулярна оси поляризации) ядра в дочерних клетках попадают в совершенно разные условия, возникшие в поляризованной цитоплазме, и вследствие этого начинают поставлять неидентичную генетическую информацию. Таким образом происходит дифференциация клеток.
Направление поляризации клеток постоянно изменяется в процессе эмбриогенеза, при образовании примордиев в апексе, в процессе закладки тканей листа, зач

