 2015-07-14
2015-07-14 1323
1323Например, стойкая мотивация жажды сопровождается возникновением в супраоптическом и в латеральном ядрах гипоталамуса длительных вспышек гипер-синхронизованной активности, которые с усилением мотивации иррадиируют на кору больших полушарий (рис. 74). Эти вспышки провоцируются также световыми и звуковыми стимулами и исчезают после внутрипо-лостного введения воды или внутривенного введения физиологического раствора. Уже эти признаки свидетельствуют о том, что при мотивации жажды в гипоталамусе формируется стойкий доминантный очаг (А. С. Батуев, Б. Г. Гафуров, 1988).
Подобного рода первичные очаги стационарного возбуждения можно рассматривать в качестве одного из системообразующих факторов при формировании доминирующей нейродинамической констелляции мозговых структур.
Анализ нейронной активности латеральных ядер гипоталамуса при формировании мотивации жажды (А. С. Батуев, Ю. Ли, 1987) обнаружил возникновение центральных и периферических перестроек во вкусовой сенсорной системе. Мотивация жажды повышает возбудимость входных рецепторных элементов вкусовой сенсорной системы и облегчает проведение к центральным нейронам информации о химических качествах пищи путем раскрытия рецептивных полей нейронов и перехода их на режим широкополосного приема.
Возникновение таких первичных очагов может вести к появлению вторичных и более мощных очагов
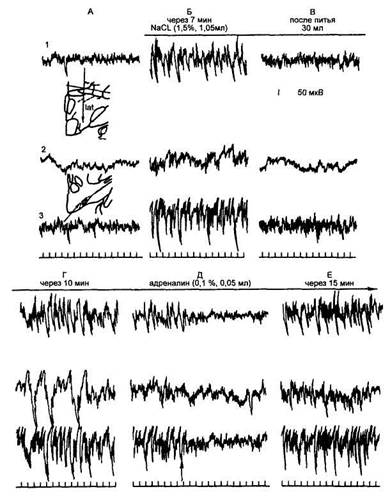
Рис. 74
Суммарная биоэлектрическая активность
коры и ядер гипоталамуса у кошек с мотивацией жажды,
созданной прямым введением хлорида натрия в латеральные ядра
гипоталамуса (по Б. Г. Гафурову и И. В. Якунину, 1989)
А — исходный фон; Б — после однократного введения хлорида натрия (горизонтальная линия сверху) появляется высоковольтная активность и поисковое поведение в сторону поилки; В — после питья 30 мл воды оба признака жажды исчезают; Г — спустя 10 мин жажда восстанавливается, Д — вновь исчезает после введения в латеральный гипоталамус адреналина; Е — через 15 мин поведенческие и биоэлектрические признаки жажды восстанавливаются; 1 — латеральный отдел гипоталамуса; 2 — сенсомоторная область коры; 3 — супраоптическое ядро гипоталамуса; вертикальная стрелка — введение адреналина. Калибровка: время — 1с, амплитуда — 50 мкВ.
в других отделах мозга, где имеются предпосылки для продолжительного удержания следовых процессов. Например, в нейронах гиппокампа посттетаническая потенциа-ция длится часами или даже сутками.
Другой структурой, где могут формироваться вторичные очаги стационарного возбуждения, является кора головного мозга. Принципиальным отличием таких очагов от первичных является исходная причина их возникновения. Так, если первичный очаг в гипоталамусе возникает и поддерживается в первую очередь за счет гуморальных факторов, то вторичные очаги в коре больших полушарий являются проекционными по своему происхождению и поддерживаются за счет широкой конвергенции к ним множества суммирующих здесь сенсорных влияний. По-видимому, следовые процессы здесь настолько мощны, что приводят к каким-то структурным изменениям, длительно модифицирующим передачу в синапсах. Одним из возможных механизмов являются стационарные изменения цитоскелета в аксошипиковых синапсах.
Если применительно к биологическим мотивациям локализацию первичных очагов можно связывать с ги-поталамическими ядрами, то топография вторичных очагов должна опираться на карту гипоталамо-кортикаль-ных связей. Р. А. Павлыгина (1982) считает, что в нормально функционирующем организме должны быть заложены механизмы, допускающие сосуществование нескольких доминант одновременно. Сопряженные тормозные взаимовлияния характерны для функционально несовместимых деятельностей, взаимоотношения между которыми закреплены генетически. Образование же рецептивного поля доминанты, то есть избрание адекватных для себя сигнальных раздражений, способных к воспроизведению и последующему поддержанию доминанты, отражает наиболее пластическое звено доминанты, также опирающееся на процессы центрального торможения. Значит, торможение как в части афферентного обеспечения доминанты, так и в части ее координированного эфферентного выполнения играет определяющую роль, и сопряженное торможение первично выступает в роли организатора суммации в наличном очаге возбуждения.
Таким образом, для обеспечения суммации сенсорных воздействий в пределах рецептивного поля кортикального вторичного «очага» повышенной возбудимости необходимо допустить торможение как второстепенных сигнализаций, так и конкурентных рефлекторных актов.
Доминанта и условный рефлекс. В своей работе «Об условно-отраженном действии» А, А. Ухтомский писал: «Шеррингтоновские принципы отмечают особо, как успело отпечатлеться уже морфологически преобладание рецептивных приборов над исполнительными (двигательными). Но впервые И. П. Павлов со своими учениками осветил тот механизм и принцип, которым это преобладание формируется на ходу работы организма. Это механизм „условного рефлекса" и принцип „временной связи". Если воронка Шеррингтона имела в виду постоянные рефлекторные дуги, закрепившие функциональную и морфологическую связи между собой наследственно и филогенетически, то И. П. Павлов улавливал самое закладывание и новообразование связи в этой воронке, привлечение все новых рецептивных сфер конвергенции относительно эфферентных (исполнительных) приборов... Связь исполнительного органа с новыми рецептивными поводами может закладываться, так сказать, случайно, вследствие того, что работа данного исполнительного органа имела случаи — более или менее совпадать во времени с данной дальней рецепцией. Кора бдительно примечает такие совпадения, закладывает по их поводу новые связи, сначала временные, а затем могущие закрепиться в качестве опыта и фонда, который будет использован животным для дальнейшей жизни... перед нами новая рефлекторная дуга в процессе своего закладывания и дальнейшего закрепления»1.
Рецептивное поле доминанты создает спектр тех сиг-нально значимых факторов среды, которые обеспечивают данную доминанту на разных стадиях ее существования. А. А. Ухтомский (1966) выделил три стадии доминанты: 1. Стадия доминанты, безразлично привлекающей к себе в качестве поводов возбуждения всевозможные рецеп-
1 Ухтомский А. А. Собр. соч. Л., 1954. Т. 4. С. 225-226.
ции. 2. Стадия образования условного рефлекса, когда из множества действующих рецепций доминанта вылавливает группу рецепций, которая для нее в особенности биологически интересна (выработка адекватного раздражителя для данной доминанты). 3. Стадия более или менее исключительной связи данной доминанты с данным рецептивным содержанием, когда каждый из контрагентов (внутреннее содержание и внешний образ) будут вызывать и подкреплять исключительно друг друга.
На первой стадии достаточно устойчивой доминанты ее рецептивное поле оказывается чрезвычайно широким: разнообразные афферентации подкрепляют наличную доминанту. Причем уже на этой стадии обнаруживается координирующая роль торможения, препятствующего выполнению иных форм деятельности (Р. А. Павлыги-на, 1982, 1988).
Признавая за доминантой способность подкрепляться разнообразными факторами среды, следует иметь в виду существование механизмов избирательного реагирования, то есть наиболее эффективных агентов среды, жестко связанных с поддержанием и реализацией данного доминантного состояния. К такого рода сигналам в первую очередь должны быть отнесены натуральные раздражители (по павловской терминологии) или ключевые раздражители (по терминологии этологов). Причем совсем не обязательно, чтобы такие свойства сигналов были бы генетически детерминированы.
Например, в определенных условиях одиночные коммуникационные сигналы животных могут приобретать характер ключевых раздражителей. Таким условием для кошек может быть возникновение биологической доминанты, проявляющейся во время лактации (Н. Н. Соколова, 1984). В стрессорных ситуациях котята издают сигналы, близкие по структуре к сигналам новорожденных. Такие сигналы неизменно вызывают реакцию материнского ухаживания у самок. Если же такие сигналы предъявляются с магнитофона, то у лактирующих самок проявляется лишь ориентировочная реакция без активного поиска котенка. Следовательно, для возникновения адекватных ответных реакций кошки-матери необходимо комплексное воздействие разномодального
сигнала, звуковые же компоненты являются лишь частью демонстрационного поведенческого комплекса, недостаточной для полного воспроизведения адекватных реакций.
Таким образом, рецептивное поле доминанты материнского ухаживания у лактирующих кошек представляет собой динамическую систему, в состав которой входят жесткие компоненты (ключевые сигналы) и пластические компоненты, биологическая значимость которых для данной доминанты оценивается в контексте всей окружающей ситуации.
Фронтальная кора у кошек-самок к моменту рождения детенышей оказывается избирательно настроенной к тем звуковым частотам, которые являются формант-ными для видоспецифических вокализаций детенышей первого месяца жизни (Г. А. Куликов, 1989). Значит, состояние доминанты обеспечивает выделение жестких компонентов (ключевых), связанных с восприятием наиболее значимой в биологическом отношении сигнализации и выполнением соответствующих жестко запрограммированных поведенческих актов.
Закономерности избирательного реагирования на сиг-нально значимые агенты среды могут служить физиологическими предпосылками для трактовки механизмов сенсорного внимания. В самом общем смысле внимание рассматривается как один из механизмов устранения избыточности сенсорных сообщений, который участвует как в избирательном подавлении, обеспечении сенсорных входов, так и в фильтрации информации, извлекаемой из систем кратко- и долговременной памяти.
Закрепление рецептивного поля с выделением адекватных для данной доминанты агентов внешней среды осуществляется по механизму условного рефлекса, в котором важнейшую роль отводят фактору подкрепления. Подкрепление условного рефлекса направлено к удовлетворению текущей доминирующей мотивации, к снижению возбудимости первичного доминантного очага. Мотивационное поведение животных направлено прежде всего на снижение доминанты. Полное удовлетворение биологической потребности, достигнутое в результате подкрепления систем условных рефлексов, при-
водит к исчезновению доминантных очагов и торможению соответствующих условных рефлексов.
Теперь обратимся к рассмотрению эндогенно вызванного доминантного состояния (например, жажды) и его условнорефлекторного удовлетворения (И. П. Никитина, 1983).
Мотивация жажды создавалась добавлением соли в пищевой рацион и ограничением воды. У животных на разные условные сигналы вырабатывались побежки к двум разным кормушкам, в одной из которых подавалась пресная, а в другой — соленая пища. При избыточном поступлении соли в организм возникал отказ животного даже от минимально соленой пищи и прекращение побежек по сигналу «соленого». В результате солевой перегрузки активируется система неспецифической оценки содержания соли в предлагаемой животному пище с последующим формированием отрицательной реакции на эту пищу. Это состояние является стойким и гетеросенсорным по своему обеспечению, ибо не только конкретный условный сигнал, но и вся окружающая ситуация его подкрепляла, то есть оно характеризовалось всеми свойствами доминантного состояния. Доминирующая мотивация жажды выступает как фактор неспецифической инициации реакций отверга-ния всего соленого и предпочтения всего пресного. Одновременно с этим могут возникнуть и реакции неспецифического поиска предмета удовлетворения доминирующей мотивации. Даже однократное положительное завершение такого поведенческого акта, санкционируемое соответствующим эмоциональным сопровождением, может закрепиться в памяти.
Значение сигнальных факторов окружающей среды выступает ярко при использовании различных приемов противопоставления условных сигналов «пресного», которые теперь подкрепляются пищей разной степени солености. По сигналам «пресного» животное направляется к соответствующей кормушке и поедает из нее пищу такой степени солености, которая ранее стойко отвергалась. Такое условнорефлекторное «опреснение» пищи (психическое опреснение) может служить удачной моделью для исследования вкусовых иллюзий.
Вероятно, пищевой условный сигнал, как и доминирующая мотивация, управляют прежде всего вкусовыми порогами, поэтому животное и поедает неадекватную сигналу пищу. Условный сигнал приобретает свойства создавать определенный диапазон вкусовых порогов, в пределах которого данная мотивация может быть реализована с наибольшей вероятностью. Висцеральная сигнализация, формирующая доминирующую мотивацию, также определяет тот пороговый диапазон вкусовых сигналов, которые способны удовлетворить биологическую потребность.
Третья стадия доминанты строится на законе «обратных отношений» А. А. Ухтомского: симптомокомплекс доминанты может репродуцироваться при наличии адекватных сигналов. Такие ретенционные способности принадлежат коре больших полушарий, которая а) формирует адекватные стимулы для доминанты, б) запускает соответствующие доминанты по поводу этих стимулов и в) обеспечивает взаимодействие и переключение доминант.
На этой стадии речь должна идти не столько об элементарных биологических доминантах (голод, жажда, половое чувство), сколько о сложном памятном следе, носящем ситуативный характер (интегральный образ), а также о динамической констелляции мозговых структур, обеспечивающих целенаправленное поведение организма.
Старая доминанта возобновляется или для того, чтобы при новых данных обойтись при помощи старого опыта, или для того, чтобы по новым данным переинтегрировать старый опыт (А. А. Ухтомский, 1950). Значит, пространственно разнесенные компоненты доминирующей констелляции представляют собой центральную модель поведенческого акта, которая сформирована по закону доминанты и условного рефлекса и записана в долговременной памяти.
Справедливо заключение П. В. Симонова, который пишет: «Комбинация (точнее — синтез) механизма доминанты с механизмами формирования условного рефлекса обеспечивают оба фактора, необходимых и достаточных для организации целенаправленного поведения:
его активный творческий характер (доминанта) и точное соответствие объективной реальности (упроченный, тонко специализированный условный рефлекс)».1
Таким образом, доминанта и условный рефлекс рассматриваются в качестве основных принципов организации целенаправленного и адаптивного поведения.
§ 43. ВЫСШИЕ ИНТЕГРАТИВНЫЕ СИСТЕМЫ МОЗГА
РЕТИКУЛО-СТВОЛОВЫЙ УРОВЕНЬ ИНТЕГРАЦИИ
Ретикулярная формация — это филогенетически старая система мозга, которая не представляет собой единого анатомического целого и морфологически является гетерогенным образованием. Трудно определить и анатомическую протяженность ретикулярной формации, которая занимает центральное положение на всем протяжении ствола (рис. 75).
1 Симонов П. В. Взаимодействие доминанты и условного рефлекса как функциональная единица организации поведения // Успехи фи-зиол. наук. 1983. Т. 14. № 1. С. 18-19.
Нейрональной основой интегрирующей функции ретикулярной формации служат длинноотростчатые клетки, аксоны которых могут распространять влияние как в каудальном, так и в ростральном направлении. Ретикулярные клетки соединяют разные отделы внутри ретикулярной формации, а последнюю со спинным мозгом, большими полушариями и мозжечком.
Каждая сенсорная система направляет пути в ретикулярную формацию. Описаны мощные влияния соматических и висцеральных нервов, что позволило считать одной из функций ретикулярной формации висцеро-соматическую интеграцию. Показаны пути в ретикулярную формацию от ядер тройничного нерва, вестибулярных ядер, верхней оливы.
Кора мозга оказывает регулирующие влияния на деятельность ретикулярной формации. Пути из ретикулярной формации в нисходящем направлении — к спинному мозгу, мозжечку, ядрам специфических систем и в восходящем — к структурам больших полушарий вплоть до коры также построены по топическому принципу, что свидетельствует о функциональной специализации ретикулярных ядер.
Для активности ретикулярных нейронов характерна аутогенная ритмика, которая служит для осуществления тонических влияний на спинной мозг и большие полушария головного мозга. Сами ретикулярные нейроны способны отвечать на раздражения практически всех афферентных путей или центральных мозговых структур. Такая обширная конвергенция влияний у разных ретикулярных единиц значительно варьирует и зависит от функционального состояния мозга. Если ретикулярный нейрон дифференцирует входные влияния по их модальностям, то полисинаптические цепи нейронов с короткими аксонами, по которым активность распространяется внутри самой ретикулярной формации, лишают первоначальный сигнал его модальной специфичности. Поэтому к ядрам таламуса поступает активация, являющаяся продуктом полимодального интегрирования на ретикулярных нейронах.
Дифференцированный характер конвергенции, топическое фракционирование и гетерогенность в организа-
ции афферентных и эфферентных связей позволяют заключить, что функции, приписываемые ретикулярной формации, касаются главным образом интеграции и ре гуляции разных видов деятельности.
П. К. Анохин (1968) обращал внимание на гетерохими-ческую чувствительность отдельных синапсов и целых по-линейрональных цепей, активируемых раздражениями различного биологического качества (боль, голод и др.).
Ретикулярные влияния по соответствующим путям могут достигать различных релейных ядер сенсорных систем и дифференцирование модулировать афферентные потоки. Показано, например, что концентрация внимания животного на зрительном объекте приводит к блокированию передачи сенсорных импульсов в слуховой системе (П. Эрнандец-Пеон, 1962). Явления, именуемые привыканием, связывают с регулирующей функцией ретикулярной формации. Поскольку привыкание рассматривают как простейший вид обучения, то ретикулярной формации приписывают известную роль в формировании условных рефлексов (А. Гасто, И. Иошии и др.).
Интегрированные на ретикулярном уровне размодаль-ные афферентные посылки с учетом мотивационных факторов формируют импульсные тонические влияния к специфическим сенсорным системам, приводящие к модуляции проведения по ним соответствующих сигнализаций. В результате создаются предпосылки для успешного синтеза различных сенсорных сигнализаций на более высоких уровнях мозга. Другая сторона восходящих ретикулярных влияний может заключаться в создании на таламокортикальном уровне такого функционального состояния (или центрального тонуса мозга), который обеспечит адекватные условия для формирования разномодального афферентного синтеза.
ТАЛАМОКОРТИКАЛЬНЫЙ УРОВЕНЬ ИНТЕГРАЦИИ
Нейрофизиология ассоциативных систем мозга. Наряду со специфическими и неспецифическими системами принято выделять в качестве самостоятельной категории ассоциативные таламокортикалъные системы. Применительно к высшим млекопитающим это своеобразные
структуры, не принадлежащие какой-либо одной сенсорной системе, но получающие информацию от нескольких сенсорных систем. Ассоциативные ядра таламуса относятся к «внутренним ядрам», афферентные входы к которым идут не от сенсорных лемнисковых путей, а от их релейных образований. В свою очередь, эти ядра проецируются на ограниченные корковые территории, которые именуются ассоциативными полями.
Согласно анатомическим данным выделяют две высшие ассоциативные системы мозга. Первая включает заднюю группу ассоциативных ядер, проецирующихся на теменную область коры, и именуется таламопарие-тальной системой. Вторая на таламическом уровне состоит из медиодорзального ядра с его проекцией на лобную область коры и называется таламофронталъной системой. Обе ассоциативные системы — продукт прогрессивной дифференциации неспецифического таламуса и достигают значительных размеров у приматов и человека.
Таламопариетальная система. Теменная кора является местом широкой гетеросенсорной конвергенции по волокнам от специфических, ассоциативных и неспецифических ядер таламуса, а также по путям от сенсорных корковых зон и симметричной коры противоположного полушария.
Световые воздействия оказывают наиболее сильный эффект на теменную кору: здесь описаны наряду с муль-тисенсорными и собственно зрительные нейроны, которые отвечают на перемещения в определенном направлении сложных геометрических фигур. Иногда эти же клетки отвечают и на звуковые воздействия, и на движения глаз.
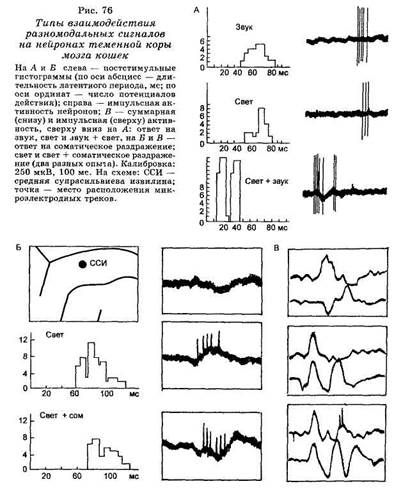
Обнаружено три типа межсенсорного взаимодействия на нейронах теменной коры: I — суммирующий нейрон, который при одновременном гетеросенсорном раздражении отвечает более сильным разрядом с более коротким латентным периодом, чем на мономодальные стимулы; И — тормозящийся нейрон, латентный период ответа которого увеличивается, когда стимул из моносенсорного превращается в гетеросенсорный, и III — детектор комплекса, который отвечает импульсным разрядом толь-
ко при комплексном гетеросенсорном воздействии и не реагирует на изолированное применение мономодальных стимулов (рис. 76). Показано при этом, что в теменной коре существуют полисенсорные клетки, отражающие докорковый, таламический уровень интеграции, наряду с такими нейронами, на мембране которых реализуются собственно кортикальные механизмы межсенсорного синтеза.
Теменная ассоциативная кора посылает мощные нисходящие связи ко многим сенсорным, лимбическим, ретикулярным и двигательным аппаратам мозга и даже образует волокна в составе кортикоспинального тракта (О. С. Адрианов, 1976).
Все вышесказанное дает основание рассматривать теменные области коры как важнейший дискриминаци-онно-интегративный аппарат больших полушарий.
После удаления теменных полей коры возникают глубокие нарушения в условнорефлекторной деятельности как на простые мономодальные, так в особенности на разномодальные комплексные сигналы. Это связано со специальной ролью теменной коры в управлении процессами селективного внимания, с оптимизацией режима текущей деятельности, а также с формированием ори-ентационных движений к опознаваемому сигналу.
В целом таламопариетальная ассоциативная система мозга представляет собой: 1) центральный аппарат первичного симультанного анализа и синтеза обстановочной афферентации и запуска механизмов ориентацион-ных движений, 2) один из центральных аппаратов «схемы тела» и сенсорного контроля текущей двигательной активности и 3) важнейший элемент долобной предпусковой интеграции, участвующий в формировании целостных полимодальных образов.
Таламофронтальная система. Сенсомоторная кора получает множественные проекции из специфических, ассоциативных и неспецифических отделов таламуса, ассоциативные кортико-кортикальные и транскаллозаль-ные входы из противоположного полушария и характеризуется наличием сложных синаптических комплексов для конвергенции на одном и том же нейроне множества афферентных влияний. Не только вставочные нейроны, но и выходные элементы сенсомоторной коры — нейроны пирамидного тракта — относятся к категории мультисенсорных клеток.
Установлено три типа ответных реакций мультисенсорных нейронов сенсомоторной коры: 1) привыкание, заключающееся в снижении вероятности ответа на каждый последующий стимул в серии; 2) сенситизация — усиление реакции при повторных применениях стиму-
ла и 3) экстраполяция — формирование упреждающего ответа нейрона на каждый последующий стимул в серии.
Такие пластические перестройки ответной активности нейронов в зависимости от характера стимуляции и функционального состояния, интенсивности и модального состава стимулов свидетельствуют о наличии сложных механизмов функциональной конвергенции, имеющих непосредственное отношение к формированию системных реакций целостного организма.
Этого нельзя сказать по отношению к собственно лобным полям на дорзолатеральной поверхности прореаль-ной извилины кошек, которые по нейрофизиологическим и морфологическим данным еще не имеют непосредственной увязки с интегративными процессами мозга, а скорее относятся к структурам неспецифического типа. В то же время медиобазальные отделы лобной коры формируют нисходящие пути к таламическим ядрам лимбичес-кой системы. Последствия удаления прореальной извилины у кошек состоят в дефиците зрительного опознания (Л. В. Черенкова, 1975) или в нарушениях точности зрительно контролируемых двигательных актов (Ю. А. Юна-тов, 1981). Последнее связывают с тем, что основное пространство собственно лобной коры кошек занимает глазодвигательное поле. Нарушений в других сенсорных системах или эмоционально-мотивационной сферы не отмечалось.
Таким образом, можно выделить следующие основные механизмы работы ассоциативных систем мозга (А. С. Батуев, 1979, 1981, 1984).
1. Механизм мультисенсорнои конвергенции. Его специфичность определяется тем, что к ассоциативным полям коры конвергируют афферентные посылки, несущие информацию о биологической значимости того или иного сигнала. Такие отселектированные афферентные влияния вступают в интеграцию на кортикальном уровне для формирования программы целенаправленного поведенческого акта.
2. Механизм пластических перестроек при гетеро-модалъных сенсорных воздействиях. Динамический характер мультисенсорнои конвергенции может проявляться
либо в избирательном привыкании, либо в сенситизации, либо наконец, в формировании экстраполяционного типа ответов. Установлен факт важной роли доминирующей мотивации в определении спектра конвергирующих модальностей и в организации внутрикорковых интеграции. Для теменной коры преобладающим может быть горизонтальный (ламинарный) тип межнейронной интеграции, а для сенсомоторной ассоциативной коры преобладающим является вертикальный (модульный) тип межнейронной интеграции.
3. Механизм краткосрочного хранения следов инте грации, заключающийся в длительной внутрикорковой или таламокорковой реверберации импульсных потоков (см. гл. 8). Последнее объясняет дефекты памяти и обучения у кошек и собак после разрушения ассоциативных полей коры или соответствующих ядер таламуса.
ЭВОЛЮЦИЯ АССОЦИАТИВНЫХ СИСТЕМ
В параллельных рядах, которыми шло развитие современных млекопитающих, хотя и сохранился общий план конструкции мозга, но таламокортикальные его системы претерпели наиболее существенные морфофункцио-нальные перестройки. Высокого развития достигают корковые механизмы деятельности сенсорных систем с ясно выраженной тенденцией возрастания ассоциативных систем мозга со свойствами полисенсорного конвергирования. В динамике морфологических преобразований происходит обособление зон перекрытия корковых проекций, с которыми связывают реализацию наиболее сложных форм высшей нервной деятельности.
В пределах класса млекопитающих можно выделить три основных уровня эволюции ассоциативных систем мозга. При этом необходимо иметь в виду, что степень развития ассоциативных формаций мозга рассматривается как показатель филогенетического статуса вида (Г. И. Поляков, 1964). Причем мозг насекомоядных рассматривается как предшественник с его дальнейшим усложнением в параллельных рядах грызунов, хищных и приматов. Исследования обнаружили нечеткость границ дифференциации внутри новой коры и неопределенность
функциональной идентификации ее полей. Это согласуется с отсутствием в таламусе четких границ между сенсорными ядрами.
Вместе с этим определенная область коры ежей (рис. 77) по морфологическим критериям позволяет допустить присущие ей интегративные свойства даже в пределах столь примитивно организованного неокортек-са (Г. П. Демьяненко, 1977). К этой области коры ежей проецируются примитивные ассоциативные ядра тала-муса — медиодорзальное и заднелатеральное. В данной области обнаружены как развомодальные моносенсорные, так и полисенсорные нейроны (А. А. Пирогов, 1977). Такие корковые элементы активируются преимущественно по единому каналу, берущему начало в недифференцированной задней группе ядер таламуса. Таким образом, в этой таламокортикальной системе сочетаются свойства как неспецифических, так и ассоциативных систем млекопитающих (рис. 77).

Было показано (И. В. Малюкова, 1974), что такая форма зрительно-слуховой интеграции, как двигательный условный рефлекс на одновременный комплекс, не может быть выработана у ежей и попытка ее сформирования приводит к невротическим срывам. Однако сложный цепной двигательный условный рефлекс может быть легко образован на мономодальный сигнал и разрушается после удаления ассоциативной области неокортекса. Больше страдают самые тонкие компоненты сенсомотор-ной интеграции, которыми завершается пищедобыватель-ный двигательный акт.
Следовательно, хотя примитивная ассоциативная система ежей еще не способна к организации сложных актов межсенсорной интеграции, но она уже начинает участвовать в осуществлении процессов сенсомоторного синтеза.

