 2015-10-16
2015-10-16 8511
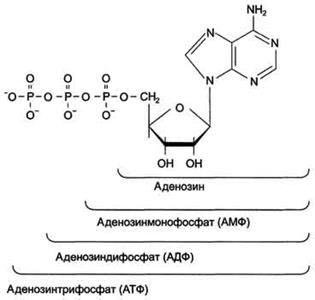
8511Каждый нуклеотид содержит 3 химически различных компонента:
1) гетероциклическое азотистое основание,
2) моносахарид (пентозу)
В зависимости от числа имеющихся в молекуле остатков фосфорной кислоты различают нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты (НДФ), нуклео-зидтрифосфаты (НТФ)
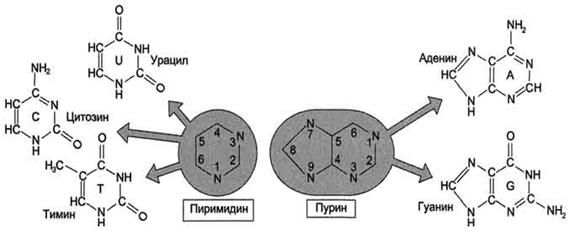
В состав нуклеиновых кислот входят азотистые основания двух типов:
1) пуриновые - аденин (А), гуанин (G)
2) пиримидиновые - цитозин (С), тимин (Т) и урацил (U).
Номенклатура нуклеотидов
| Азотистое основание | Нуклеозид | Нуклеотид | Трехбуквенное обозначение | Однобуквенный код |
| Аденин | Аденозин | Аденозинмонофосфат | АМФ | А |
| Гуанин | Гуанозин | Гуанозинмонофосфат | ГМФ | G |
| Цитозин | Цитидин | Цитидин монофосфат | ЦМФ | С |
| Урацил | Уридин | Уридинмонофосфат | УМФ | U |
| Тимин | Тимидин | Тимидинмонофосфат | ТМФ | Т |
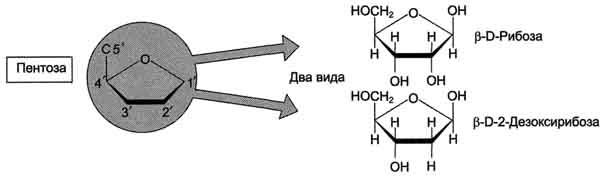
В молекулы РНК входят аденин (А), урацил (U), гуанин (G) и цитозин (С),
В молекулы ДНК - аденин (А), тимин (Т), гуанин (G) и цитозин (С).
Структура дезоксирибонуклеиновой кислоты (ДНК):
Первичная структура ДНК -порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинуклеотидной цепи.
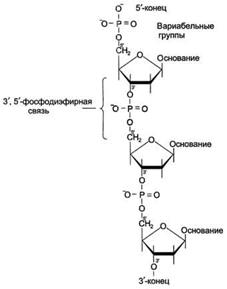
Связь между мономерами обозначают 3', 5'-фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращённо записывают с помощью однобуквенного кода, например -A-G-C-T-T-A-C-A- от 5'- к 3'-концу.
Вторичная структура ДНК. В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотиднье цепи в ней антипараллельны, т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи.
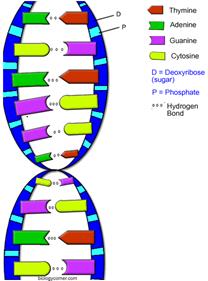
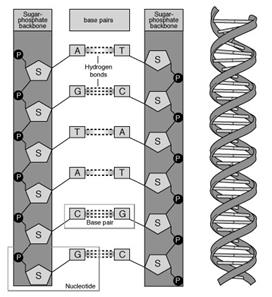
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи). При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль. Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. Длина одного витка 3,37 нм. В образованной структуре различают две бороздки - большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.








