 2015-10-16
2015-10-16 3127
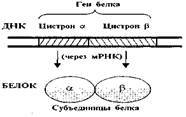
3127Информация о структуре белков и РНК записана в участках ДНК, называемых генами и цистронами.
Ген - это участок ДНК, кодирующий один белок.
Цистрон же - участок ДНК, кодирующий одну полипептидную цепь.
Таким образом, если белок состоит из нескольких разных полипептидных цепей (субъединиц), то его ген включает несколько цистронов (рис. 1).
Однако такое подразделение относится, в основном, к бактериям, где цистроны одного гена обычно следуют на ДНК друг за другом.
У животных же и человека цистроны нередко располагаются в разных хромосомах и обычно тоже называются генами: например, ген альфа-цепи и ген бета-цепи гемоглобина.
Рис. 1. Ген и цистрон
 Кроме генов всех белков организма, в хромосомах имеются также гены РНК. Среди них четыре вида генов для рибосомных РНК и нескольких десятков генов для транспортных РНК.
Кроме генов всех белков организма, в хромосомах имеются также гены РНК. Среди них четыре вида генов для рибосомных РНК и нескольких десятков генов для транспортных РНК.
Общая совокупность генов, определяющих наследственную информацию организма, называется геномом.
Всего в геноме бактерий (конкретно - кишечной палочки) - около 2500 цистронов. В хромосомах человека число генов около 30 000.
Почти все гены эукариот (в отличие от генов прокариот) имеют характерную особенность: содержат не только кодирующие участки - экзоны, но и некодирующие - интроны (рис. 2.). Экзоны и интроны перемежаются друг с другом, что придает гену как бы «разорванную» структуру.
Рис. 2. Экзоны и интроны.
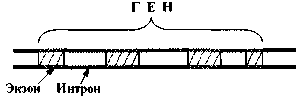 Число интронов в гене варьирует от 2 до нескольких десятков; в гене миозина их около 50. Порой на интроны приходится до 90 % общей длины гена.
Число интронов в гене варьирует от 2 до нескольких десятков; в гене миозина их около 50. Порой на интроны приходится до 90 % общей длины гена.
Между генами также находятся некодирующие последовательности - спейсеры. Несмотря на общее название, функциональная роль их может быть различной.
Многие спейсерные участки, видимо, выполняют структурную роль: - участвуют в правильной укладке нуклеосомной цепи в высшие структуры хроматина, в прикреплении хромосом к аппарату центриолей и т. д. Другие спейсеры являются специфическими местами (локусами) связывания определенных белков функционирующих на ДНК (ДНК-полимеразного комплекса и белков, выполняющих регуляторную функцию).
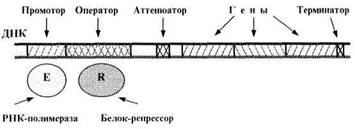
При этом участки связывания РНК-полимеразы (фермента, синтезирующего РНК на ДНК) называются промоторами. Они либо вплотную примыкают к началу гена (или группы генов), либо отделены от гена какими-либо другими функциональными локусами.
Характерный компонент промоторов у кишечной палочки - т. н. бокс (последовательность) Прибнова:
(5')-ТАТААТ-(3')
(3')-АТАТТА-(5').
Он находится за 15 нуклеотидных пар (н. п.) от стартовой точки транскрипции. Общая же протяженность промотора - несколько десятков н. п.
В случае эукариот промотор - более сложное понятие, поскольку РНК-полимераза связывается с ДНК не непосредственно, а лишь вместе с комплексом других белков - т. н. общих факторов транскрипции.
В промоторе эукариот различают небольшую область инициации, ТАТА-бокс (сходный с боксом Прибнова у бактерий) и ряд других типичных участков.
Что касается участков связывания регуляторных белков, то у бактерий их называют операторами. Они располагаются после промотора (рис. 3). При определенных условиях с оператором связывается специфический белок-репрессор, и это блокирует «прочтение» РНК-полимеразой соответствующей группы генов. Еще одним регулятором работы гена может быть - аттенюатор – это выключатель - участок ДНК между оператором и структурными генами, на котором при определенных условиях прекращается транскрипция.
Рис. 3. Функциональные отделы в ДНК бактерий.
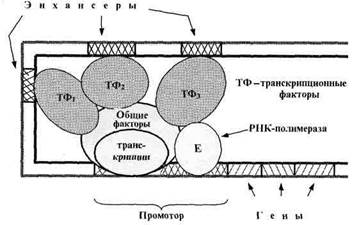
 У эукариот (включая человека) регуляцию «прочтения» генов осуществляют не только белки-репрессоры, но и белки-активаторы - транскрипционные факторы (рис. 4).
У эукариот (включая человека) регуляцию «прочтения» генов осуществляют не только белки-репрессоры, но и белки-активаторы - транскрипционные факторы (рис. 4).
К последним относятся общие факторы транскрипции, необходимые для связывания РНК-полимеразы с промотором. Эти факторы имеются во всех клетках и необходимы для «прочтения» любого функционирующего гена.
Другие транскрипционные факторы повышают активность только определенных генов. Локусы ДНК, связывающие такие факторы, называются энхансерами.
Энхансеры могут располагаться достаточно далеко от регулируемого гена: на расстоянии нескольких тысяч нуклеотидных пар.
Рис. 4. Функциональные отделы в ДНК эукариот.
 Как же связывание с ними транскрипционных факторов может стимулировать активность гена?
Как же связывание с ними транскрипционных факторов может стимулировать активность гена?
Видимо, дело в сложной пространственной организации ДНК. Она образует петли, благодаря чему энхансеры сближаются с промоторной зоной и связанные с ними факторы влияют на активность транскрипционного комплекса.
Причем, для некоторых ключевых генов в клетке имеется сразу несколько энхансеров, достаточно удаленных друг от друга. Следовательно, все они в результате изгибов ДНК должны собираться примерно в одном месте пространства.
Эта ситуация напоминает формирование активного центра в молекуле фермента. Там тоже аминокислотные остатки, образующие этот центр, расположены в разных участках полипептидной цепи и собираются вместе лишь при формировании третичной структуры.
Что же касается белков-репрессоров, то они при вхождении в состав транскрипционных комплексов не повышают, а понижают их активность.
Часто используется также термин «супрессоры» - так называются факторы (обычно это белки), которые угнетают тот или иной клеточный процесс. Например, опухолевые супрессоры - регуляторные белки, препятствующие опухолевому росту. Очевидно, по механизму своего действия они могут быть как репрессорами, так и транскрипционными факторами.
Наконец, в цепи ДНК после структурных генов могут содержаться короткие локусы - терминаторы, служащие сигналами об окончании (терминации) транскрипции ДНК.
Дополнительная литература.
1. Грин Н., Стаут У., Тейлор Д. Биология: В 3-х т. Т.3: Пер с англ. / Под ред. Р. Сопера. – М.: Мир, 1993. С.217-221.
2. Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. М.: 2003 554 с.








