 2013-12-29
2013-12-29 1346
1346Репликация ДНК у эукариот
ДНК эукарит организована в нуклеосомы и находится в составе хроматиновых волокон. Эукариотическая ДНК также реплицируется в двух направлениях, но репликативные вилки движутся в 10 раз медленнее, чем у E.coli. Чтобы процесс репликации шел быстрее, существует много точек начала репликации (число которых, вероятно, превышает тысячу). Из каждой такой точки одновременно в противоположных направлениях движутся две репликативные вилки (рис.9), благодаря чему репликация целой эукариотической хромосомы может завершиться даже быстрее, чем репликация бактериальной хромосомы. Поскольку в эукариотической клетке хромосом много, все они должны реплицироваться одновременно. Таким образом, в ядре эукариотической клетки работает одновременно много тысяч репликативных вилок.
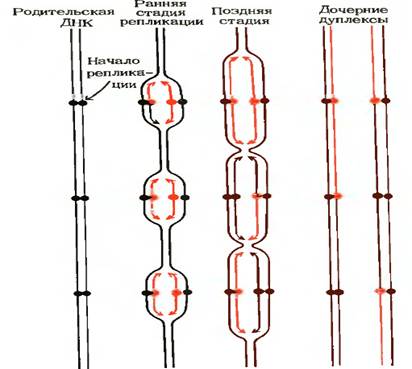
Рис. 9 Репликация эукариотической хромосомы
Рассмотрим вопрос об участии ферментов в репликации ДНК.
Фермент ДНК-полимераза I катализирует последовательное присоединение дезоксирибонуклезидных остатков к концу цепи ДНК с одновременным высвобождением неорганического пирофосата. Уравнение реакции имеет вид:
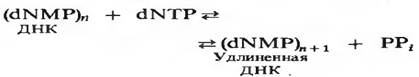
где dNMP и dNTP означают соответственно дезоксирибонуклеозид-5 -монофосфат и дезоксирибонуклеозид-5 -трифосфат. Если хотя бы один из четырех предшественников отсутствует, то новая ДНК не образуется, т.е. синтез новой ДНК идет только в присутствии всех четырех предшественников. 5 -трифосфаты всех четырех дезоксирибонуклеозидов не могут быть заменены соответствующими 5 -дифосфатами или 5 -монофосфатами. Фермент не работает также с рибонуклеозид-5 -трифосфатами. Для работы ДНК-полимеразе необходимы ионы Mg2+, а в ее активном центре содержится прочно связанный с ферментом ион Zn2+.
Синтез цепи ДНК происходит в направлении 5 → 3 (рис. 10).
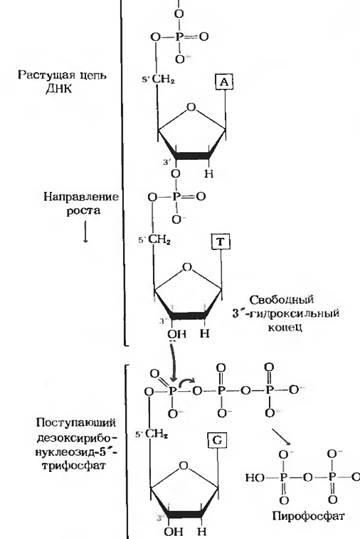
Рис. 10 Удлинение цепи ДНК с помощью ДНК-полимеразы
Энергия, затрачиваемая на образование каждой новой фосфодиэфирной связи в остове ДНК, обеспечивается расщеплением пирофосфатной связи в дезоксирибонуклеозид-5 -трифосфатах. Образующийся при этом пирофосфат разрушается затем до фосфата. ДНК-полимеразная реакция протекает только в том случае, если в системе уже находится некоторое количество предшествующей двухцепочечной ДНК.
Корнберг и его коллеги установили, что ДНК выполняет в полимеразной реакции две важнейшие функции – она служит затравкой и матрицей.
ДНК-полимераза последовательно добавляет нуклеотиды к 3 -концу одной из цепей, служащей затравкой. Следовательно, синтез новой цепи происходит в направлении 5 → 3 . ДНК-полимераза не в состоянии саамам по себе без затравки начать синтез новой ДНК. Она способна только удлинять уже существующую цепь, причем даже это она может делать только в присутствии цепи, играющей роль матрицы.
Нуклеотиды присоединяются к цепи-затравке в соответствии с нуклеотидной последовательностью цепи-матрицы по принципу комплементарного спаривания Уотсона-Крика. Где бы ни находился в матричной цепи остаток тимина, в дочерней цепи в этом месте встраивается остаток аденина, и наоборот. Аналогичным образом, если в цепи-матрице стоит остаток гуанина, то напротив него в дочерней цепи будет встроен остаток цитозина, и наоборот. Таким образом, продукт ДНК-полимеразной реакции – это дуплекс с комплементарно спаренными основаниями. На рис. 11 схематически показано, какую роль играет в ДНК-полимеразной реакции предшествующая ДНК.
Рис. 11 Структура предшествующей двухцепочечной ДНК, которая требуется для действия ДНК-полимеразы. Для того чтобы шел синтез, нужна одиночная неспаренная цепь, которая служит матрицей. Кроме того, необходимо присутствии цепи-затравки, к которой присоединяются новые нуклеотидные звенья.
Поскольку ДНК-полимеразе необходимы как цепь-затравка, так и свободная цепь-матрица, этот фермент не способен осуществлять репликацию целой нативной хромосомы, если последняя является двухцепочечным кольцом, одноцепочечным кольцом или интактным линейным дуплексом, в котором спарены все основания.
В настоящее время известно, что для репликации необходима не только ДНК-полимераза. В этом процессе участвуют больше 20 различных ферментов и белков, каждый из которых выполняет определенную функцию. Репликация состоит из большого числа последовательных этапов, которые включают узнавание точки начала репликации, расплетание родительского дуплекса, удержание его цепей на достаточном расстоянии друг от друга, инициацию синтеза новых дочерних цепей, их элонгацию, закручивание цепей в спираль и терминацию репликации. Все эти этапы процесса репликации протекают с очень высокой скоростью и исключительной точностью. Весь комплекс, состоящий более чем из 20 репликативных ферментов и факторов, называют ДНК-репликазной системой, или реплисомой.
Рассмотрим в общих чертах основные этапы процесса репликации.
Клетки E.coli содержат три различных ДНК-полимеразы, обозначаемые I, II, III (табл. 3).
Таблица 3 Характеристики ДНК-полимераз E.coli
В интактных клетках E.coli за элонгацию цепи ДНК отвечает главным образом ДНК-полимераза III. Она функционирует в форме большого комплекса с молекулярной массой ~ 550 000, называемого холоферментом ДНК-полимеразы III (рис. 12). Этот комплекс содержит в своем составе ион Zn2+ и требует для работы наличия ионов Mg2+. Ему также необходима матрица и затравка, без которой он не может инициировать процесс репликации. Подобно ДНК-полимеразе I, он удлиняет ДНК направлении 5 → 3 , присоединяя новые нуклеотиды к 3 -концу цепи-затравки.
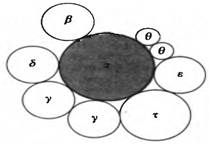
Рис. 12 Холофермент ДНК-полимеразы III
Оба фермента – ДНК-полимераза I и ДНК-полимераза III – обладают тремя ферментативными активностями. Кроме полимеразной активности они имеют 5 → 3 и 3 → 5 - экзонуклеазные активности, т.е. они могут отщеплять концевые нуклеотиды с любого конца цепи ДНК. Функция ДНК-полимеразы II пока неизвестна.
Японский ученый Р.Оказаки обнаружил, что в ходе репликации ДНК E.coli и других бактерий большая часть новообразованной ДНК обнаруживается в форме небольших кусков. Эти куски, получившие название фрагментов Оказаки, содержат приблизительно 1000-2000 нуклеотидных остатков у прокариот и 150-200 нуклеотидов у эукариот. Оказаки предположил, что эти фрагменты представляют собой короткие отрезки молекулы ДНК, образующиеся в результате прерывистой репликации и затем соединяющиеся друг с другом в одну цепь. Благодаря этому открытию удалось показать, что одна из цепей ДНК реплицируется непрерывно в направлении 5 → 3 , т.е. в направлении движения репликативной вилки; эту цепь называют ведущей. Другая цепь синтезируется прерывисто с образованием коротких фрагментов также за счет присоединения новых мономеров к 3 - концу, т.е. в направлении, противоположном движению репликативной вилки. Затем фрагменты Оказаки с помощью ферментов сшиваются друг с другом, образуя вторую дочернюю цепь, называемую отстающей (рис. 13).
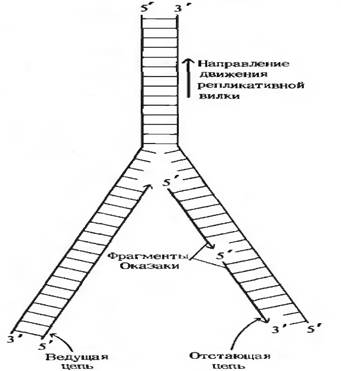
Рис. 13 Прерывистая репликация одной из цепей ДНК
Для синтеза фрагментов Оказаки в качестве затравок требуются короткие отрезки РНК, комплементарные матричной цепи ДНК. Эта РНК образуется в направлении 5 → 3 из АТФ, ГТФ, ЦТФ и УТФ с помощью фермента, называемого примазой. К 3 -концу этой короткой одноцепоченой РНК – затравки присоединяются друг за другом дезоксирибонуклеотидные остатки, комплементарные цепи-матрице ДНК. Обычно РНК-затравка состоит всего лишь из нескольких рибонуклеотидных остатков, к которым затем ДНК-полимераза III присоединяет 1000-2000 дезоксирибонуклеотидных остатков, и в результате образуется фрагмент Оказаки. Естественно, нуклеотидная последовательность новосинтезированного фрагмента Оказаки комплементарна нуклеотидной последовательности соответствующего участка цепи-матрицы. После завершения синтеза фрагмента Оказаки РНК – затравка удаляется, нуклеотид за нуклеотидом, с помощью 5 → 3 - экзонуклеазной активности ДНК- полимеразы I. Новый фрагмент Оказаки присоединяется к отстающей цепи ДНК с помощью фермента ДНК-лигазы.
Двойная спираль ДНК представляет собой плотно скрученную структуру. Кодирующие основания находятся внутри спирали, чтобы реплицирующие ДНК-ферменты смогли «прочитать» нуклеотидную последовательность матрицы, цепи родительской ДНК должны быть разделены хотя бы на коротком участке.
Раскручивание двойной спирали и удержание двух цепей на некотором расстоянии друг от друга, чтобы они могли реплицироваться, осуществляется нескольких специальных белков (рис. 14).
Ферменты, известные под названием хеликаз, расплетают короткие участки ДНК, находящиеся непосредственно перед репликативной вилкой. Для раскручивания ДНК требуется энергия. На разделение каждой пары оснований расходуется энергия гидролиза 2-х молекул АТФ до АДФ и фосфата. Как только небольшой участков ДНК оказывается расплетены, к каждой из разделяющих цепей прочно присоединяются несколько молекул ДНК-связывающего белка (ДСБ), который препятствуют образованию комплементарных пар и обратному воссоединению цепей. Благодаря этому нуклеотидные последовательности цепей ДНК оказываются доступными для репликативной системы. ДНК- полимераза может непосредственно удлинять ведущую цепь, добавляя к ее 3 -концу новые нуклеотиды. Другие специфические белки помогают примазе получить доступ к матрице для отстающей цепи. В результате примаза получает возможность связываться с отстающей цепью и синтезировать РНК-затравки для фрагментов Оказаки.
Быстрое раскручивание цепей родительской ДНК в процессе репликации (4500 об/мин) порождает еще одну проблему, которая состоит в том, что при отсутсвии специального шарнирного устройства вся хромосома, расположенная впереди репликативной вилки, должна вращаться с такой же скоростью. Предполагают, что избежать этого помогает клетке шарнир в ДНК (прямо перед репликативной вилкой), благодаря которому вращаться с большой скоростью приходится только короткому участку ДНК. Это может быть достигнуто за счет кратковременного разрыва одной из цепей ДНК, который очень быстро и точно восстанавливается после одного или нескольких оборотов. Кратковременные разрывы и воссоединения осуществляются ферментами топоизомеразами. У прокариот топоизомераза называется ДНК-гираза.
Рис. 14 Схематическое изображение основных этапов репликации ДНК
По мере того как репликативная система ликвидирует разрывы в отстающей цепи, она движется вдоль реплицирующейся ДНК. Две новые цепи соединяются со своими комплементарными цепями-матрицами автоматически, образуя две дочерние спирали, каждая из которых содержит одну родительскую и одну новосинтезированную цепь. Для образования новых спиралей не требуется ни затрат энергии, ни участия какого-либо «закручивающего» фермента.
Механизм действия ДНК-полимераз эукариот подобен таковому у прокариот. Линейность ДНК эукариот является причиной проблем, которых не существует у прокариот, имеющих кольцевую ДНК. В отличие от лидирующей цепи, которая реплицируется непрерывно, праймер (отрезок РНК), находящийся у 3 - конца отстающей цепи, разрушается и не реплицируется при помощи ДНК-полимераз. Для предотвращения укорачивания цепи на концах хромосомы находятся теломеры - участки нереплицируемой ДНК. На этом участке ДНК может синтезироваться праймер, и полнота репликации сохранится. Теломера состоит из большого числа повторов, например, у человека: ТТАГГГ. Матрицей для теломеры является РНК, а специальный фермент теломераза, представляющий собой обратную транскриптазу, присоединяет эти фрагменты к 3 - концу для сохранения исходных размеров хромосомы.

