2020-01-14
2020-01-14 444
444-Эктопаразиты - находятся на поверхности тела хозяина (блохи, вши, клещи и др.). У них имеются специализированные приспособления, позволяющие удержаться на теле хозяина.
-Эндопаразиты - приспособились к обитанию внутри организма хозяина, в его органах, тканях, клетках.
-Временный-использует хозяина только для питания
-Постоянный-все стадии развития происходят в хозине
Пути заражения:
Перорально(через рот), перкутанно(через кожу), пермукозально(через слизистую), трансплацентарно, трансвагинально.
Способы заражения:
Трансмиссивно – с помощью спецефического переносчика.
Нетрансмиссивно – алиментарный(паразит в пище), контактный, водный, инструментальный(на гразных мед.инструментах).
Виды хозяев, путей и способов заражения.
Хозяевами паразитов могут быть:бактерии,простейшие,растения,животные,человек
Виды хозяев:
1.окончательный – в нем живет половозрел особь или происх размож параз полов путем
2.промежуточный - в нем параз живет в личин стадии и размнож беспол путем
3.прокормитель – ор-м кот служ только для пит параз
4.резервуарный – ор-м в кот происх накопл параз без его развития
Перносчики: механич (только перносят параз на лапках, щетинках), специфич (в них происх опред стадии развит параз)
резервуар – ор-м в кот параз нах-ся длит время, пит и размнож. Это источн зараж.
Способы:
Трансимс: инокулятив(через рот апп), контамитив.
Нетрансмис: алиментар, водный, контактный, трансфуз, инструмент.
Аутоинвазия – зараж-е пара, кот уже сущ в хоз-ве и начин новый цикл без выхода из хозяева. (свин цеп)
Аутореинвазия – с выходом.
3. Аскарида,острица



4. Значение популяционного метода в генетике человека.
Одним из важных направлений в современной генетике является популяционная генетика. Она изучает генетическую структуру популяций, их генофонд, взаимодействие факторов, обусловливающих постоянство и изменение генетической структуры популяций. Под популяцией в генетике понимается совокупность свободно скрещивающихся особей одного вида, занимающих определенный ареал и обладающих общим генофондом в ряду поколений. (Генофонд — это вся совокупность генов, встречающихся у особей данной популяции).
В медицинской генетике популяционно-статистический метод используется при изучении наследственных болезней населения, частоты нормальных и патологических генов, генотипов и фенотипов в популяциях различных местностей, стран и городов. Кроме того, этот метод изучает закономерности распространения наследственных болезней в разных по строению популяциях и возможность прогнозировать их частоту в последующих поколениях.
Популяционно-статистический метод используется для изучения:
а) частоты генов в популяции, включая частоту наследственных болезней;
б) закономерности мутационного процесса;
Этногеномика - шаг в исследовании человека
БИЛЕТ 40
1.РЕЦЕПТОРНО-СИГНАЛЬНАЯ ФУНКЦИЯ, МЕХАНИЗМЫ ПЕРЕДАЧИ СИГНАЛА В КЛЕТКЕ
Обеспечивает р-ии кл на изменения факторов внутр и внеш среды, в результате происходит саморегуляция клеточной жизнедеятельности. Узнавание сигнала осущ спец молекулы-рецепторы-протеины и гликопротеины ПАК. Их наз мембранными рецепторами. Они имеют рецепторный, трансмембранный и цитоплазматический домены.
-Рецепторный – главный домен, с его помощью поисходит распознание сигнала. Хеморецепторы узнают хим сигналы, фоторецепторы – световые, терморецепторы – температурные изменения, а барорецепторы – перепады давления.
-Трансмембранный домен – фиксирует рецептор БЛС и участвует в передаче сигнала на ЦП-еский домен
-ЦП домен – эффекторный.
Рецепторные мол могут располагаться на периферич гиалоплазме – ДНК-связывающий домен – позволяет им фиксироваться в ядре и участвовать в регуляции работы генов.
Передача сигнала осущ путём изм конформации рецептора, кот активизирует др клеточные Б(ответные р-ии мб быстрыми и медленными).-Быстрые – осущ двухкомпонентными рецепторно-сигнальными системами, кот состоят из мембранного рецептора и ферм протеинкиназы. -Медленные – осущ универсальными си-ми, вкл вторичных посредников, кот усиливают исходный сигнал, поступающий в кл. Обычно вкл G-белки, они исп замену ГДФ на ГТФ, чтобы активировать ферм синтеза вторичных посредников.
Сигнальными мо явл гормоны, нейромедиаторы, иммуномедиаторы,факторы роста, лек препараты. Они выступают в роли первич посредников. Стероидные гормоны проникают в кл через БЛС, а инсулин или ацетилхолин не проникают в кл, но могут связываться с мембранным рецептором, обр цепь последовательных событий в мембраны и внутри кл. Первым событием явл изм конформации ЦП-ого домена рецептора, активирующего G-белок, кот обменивает ГДФ на ГТФ, диссоциирует на 3 субъединицы: а, В, у. а – обладает ГТФ-азной активностью и расщепляет ГТФ, изменяя конформацию.
-Гуанилат-циклазная си-ма с участием цикло-ГМФ. – действует как антагонист за счёт размыкания цикло-ГМФ и цикло-АМФ.
Хим в-ва могут вызывать активацию и ингибирование Рецепторно-сигнальных с-м, а так же менять их с-ва.Ех:кофеин блокирут фосфодиэстеразную систему, о активирет аденилат-циклазнуу.
Нарушения в строении функции рецепторов приводят к развитию патологий.
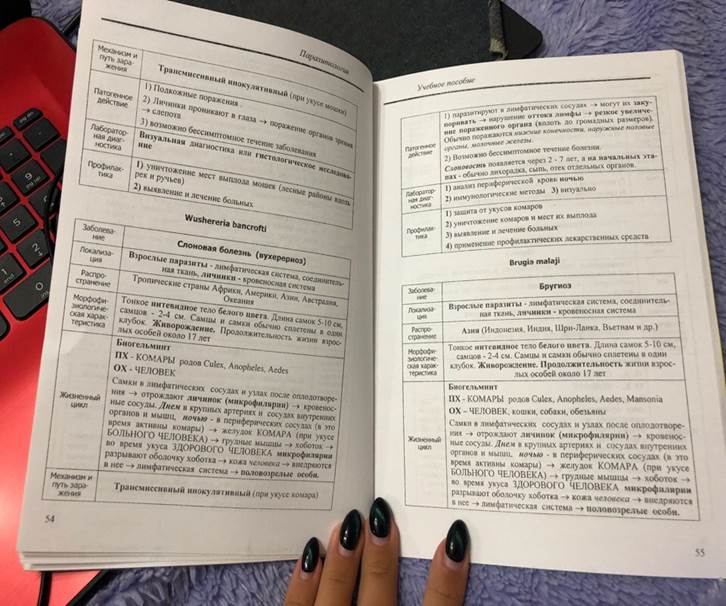
2. Филярии. Строение и жизненные циклы.
Тип – Круглые черви
Класс – Собственно круглые черви
Биогельминты


3.. Генные мутации.
Изменчивость - способность живых организмов приобретать новые признаки и свойства. Изменчивость отражает взаимосвязь организмов с внешней средой. Различают ненаследственную и наследственную изменчивость.
Ненаследственная, или фенотипическая, или модификационная, изменчивость — изменения признаков организма, не обусловленные изменением генотипа.
ГЕННЫЕ МУТАЦИИ связаны с изменением состава или последовательности нуклеотидов ДНК в пределах гена. Генные мутации наиболее важны среди всех категорий мутаций. Генные мутации возникают при изменении химической структуры гена. Это происходит в результате замены одной или нескольких пар азотистых оснований, или мутаций со сдвигом рамки считывания информации, связанных с выпадением или вставкой одного или нескольких азотистых оснований.Мутации, затрагивающие одну пару оснований и приводящие к замене на другую, удвоению, делеции, называют точковыми. Происходит нарушение последовательности нуклеотидов в молекуле ДНК. Это приводит к изменению строения белка. Генные мутации возникают при замене, выпадении, вставке пар нуклеотидов. Большинство мутаций - генные. С ними связаны изменения морфологических, биохимических, физиологических признаков.
4.. Онтофилогенетические пороки развития опорно-двигательной системы.
Врожденные пороки развития, аномалии развития, врожденные дефекты - это синонимы нарушений структуры, поведения, функций и метаболизма, встречаются в любом возрасте. Клиническое их значение и частота разные.
Они делятся на единичные и множественные, такие, что имеют легкое течение и не проявляются клинически проявления при жизни, и такие, которым свойственен тяжелый, иногда несовместим с жизнью течение. Наука, изучающая причины этих нарушений, называется тератология (от греч. τέρας - чудовище, урод). Большинство структурных аномалий у 2-3% новорожденных детей; в 2-3% детей аномалии обнаруживают в течение первых 5 лет жизнь. Врожденные дефекты являются основной причиной детской смертности. Причины врожденных пороков на 40 - 60% остаются невыясненными. Генетические факторы обусловливают примерно 15% пороков, факторы окружения - 10%, мультифакториальные наследственность - 20-25%. Тератогены, вызывающих пороки развития человека, приведены ниже. Пороки возникают в процессе формирования структур, в ходе онтогенеза. Большинство из них возникает в промежутке между третьим и восьмым неделями беременности Процессы развития (онтогенез) достаточно сложны. Так, рука у человека состоит из 29 костей и каждая по- должна иметь определенную форму, размеры, образовывать с другими костями сложные суставы. Кроме того, должно образоваться более 40 мышц определенных размеров и прикрепиться к четко определенных участков. Сюда подходит много нервных стволов, артерий, вен и т.д.. Предпосылкой возникновения врожденных пороков является действие различных факторов в критические периоды онтогенеза.
Аномалии скелета.
- Краниошиз - незавершенный процесс формирования свода черепа, что приводит к анэнцефалии.
- Скафоцефалия (ладьевидный череп) – преждевременное закрытие стреловидного шва.
- Акроцефалия (башенный череп) – преждевременное закрытие венечного шва.
- Плагиоцефалия (асимметричный краниосиностоза) - преждевременное закрытие венечного и лямбдоподибного швов только с одной стороны.
- Менингоцеле, менингоенцефалоцеле и менинго- гидроенцефалоцеле - аномалии, обусловленные нарушением окостенения костей черепа.
- Микроцефалия проявляется уменьшением свода черепа. Основанием этого является аномальное развитие мозга.
- Мандибулолицевий дизостоз характеризуется гипоплазией скул, нижней челюсти, косыми нисходящими щелями век, искажением внешних ушей.
- Гипертелоризм.
- Пивлицева микросомия - черепнолицеви аномалии верхней челюсти, височной и скуловой костей.
- Меромелия - отсутствие части конечностей.
- Амелия - полное отсутствие одного или нескольких конечностей.
- Полидактилия - сверхкомплектные пальцы кисти или стопы.
- Синдактилия - аномальное сращение пальцев кисти или стопы, щель кисти и стопы.
126. Современная синтетическая теория эволюции. Микроэволюция.
Учение об эволюции органического мира. Элементарной единицей эволюции- популяция, Механизм эволюции: случайные мутации на генетическом уровне и наследование наиболее удачных с мутаций.
1) Теория микроэволюции- совокупность процессов внутри вида. Сущность: изменение структуры популяции. В результате происходит замещение аллеля. Популяции являются открытыми генетическими системами, поэтому адаптивный признак может перейти из одной популяции в другую. Мутации и поток генов создают необходимое разнообразие генотипов популяции- то есть изменчивость. Изоляция оберегает популяцию от проникновения генетического материала извне. Естественный отбор и дрейф генов сортируют генотипы. Мутации без естественного отбора не ведут к появлению новых признаков.
Изоляция приводит к накоплению и закреплению изменений. Она бывает территориальная (географическая, экологическая и репродуктивная) или биологическая.
Дрейф генов – случайное отклонение от постоянной частоты встречаемости генов. Чем меньше популяция, тем больше вероятность случайных отклонений в ее генофонде. Для того, чтобы дрейф давал эффект, популяция должна быть хорошо изолирована. Дрейф закрепляет редкие генотипы. Все эволюционные изменения носят случайный характер, так как базируются на единичных случайных изменениях в генетическом аппарате отдельных особей.
2) Теория макроэволюции изучает происхождение надвидовых таксонов (семейств, отрядов, классов и т.д.), основные направления и закономерности развития жизни на Земле в целом, включая возникновение жизни и происхождение человека как биологического вида.
БИЛЕТ 41
1.СТРОЕНИЕ ПАК.
1)Плазмолема. Состоит из липидов(фосфолипиды, гликолипиды, стероиды, холестерин) и белки.
Фосфо- и гиколипиды-сложые эфирные соединения многоатомных спиртов-глицерина и сфингозина с жк. Липиды, сод трёхатомный спирт глицерол, наз глицеролипидами, а многоатомный спирт сфингозин-сфинголипидами.
Все молекулы липидов имеют полярное строение: гидрофильную заряженную головку, нейтральную шейку и 2 гидрофобных незаряженных хвоста. Головка может сод остаток фосфорной к-ты или углеводы, такие липиды наз фосфо- и гликолипидами. Хвосты липидов сод остаток жк. В шейке трёхуглеродный спирт-глицерол или сфингозин. За счёт полярного строения липиды обладают амфмипатичностью, благодаря кот в водн ср они способны самоорганизовываться в замкнутые пузырьки-мицеллы или БЛС. Такой вариант организации считается термодинамически устойчивым. Мол липидов: «флип-флоп» переход из одного ипидного слоя в другой; чем быстрее движутся липиды, тем более жидкостная мембрана и наоборот.) Холестерин(ол) явл природным жирным спиртом, кот выбрасывается клетками печени, кишечника и почек; в клеточных мембранах стабилизирует их плотность и регулирует проницаемость клеток. Все мембранные липиды выполняют структурную функцию, осущ барьерно-изолирующую функцию и регулируют работу мембранных белков и транспорт в-в через кл, влияя на жидкостность мембраны.
Модели строения: 1)«Бутербродная» 2 сплошных белковых слоя над и под БЛС(мембрана эритроцитов). Энергозатратна для транспорта. 2) «Липопротеиновый коврик» переплетение молекул белков и липидов между собой(МТ), очень низкая проницаемость. 3) Жидкостно-мозаичная Избирательная проницаемость, основу сост БЛС, в кот вкраплены мол мембранных белков(интегральные-цеиком или частично погружены. Трансмембранные-1 или 2 раза пересекают блс(иммуноглобулины, ферм пристеночного пищеварения) или обр канал, через кот осущ транспорт молекул. Нетрансмембранные-не пересекают блс, но ковалентно связаны с липидами; периферические-слабыми электростатическими связями).
2)Надмембранный аппарат-вкл гликокаликс и внекл структуры. ГК ввязан с плазмолемой и его компоненты представлены: -углеводными остатками гликолипидов и гикопротеинов, обр ПЛ –нек полуинтегральными и периферическими белками плазмолемы –свободными углеводами
Углеводы гликокаликса явл олигосахаридными и полисахаридными цепочками, присоединёнными к трансмембранным и периферическим белкамили липидам. Свободные углеводы представлены гликозамингликанами или мукополисахаридами.
Белковый компонент гликокаликса вкл в себя периферичесие белки, липопротеины и ацилпротеины. К белковому компоненту относят надмембр домены интегральных белков. Производными Надмембр аппарата явл некл структуры, в соста кот входят белки и гликопротеины.
3)Субмембранный опорно-сократительный аппарат – располагается под плазмолемой, из 2 частей
- Периферическая гиалоплазма это водн р-р солей, сахаров, АК и белков, явл микросредой для ОСС. Опорно-сократительная с-ма вкл в сбя тонкие фибриллы, микрофибриллы, промежут филаменты и МТ.
Тонкие фибриллы(ТФ)-тонкие белковые нити, обрфибрилярную сеть, участвуют в обр цитоскелета и связывают между собой микрофибриллы, промежут филаменты и МТ
Микрофибриллы(МФ)-тонкие и толсты нитевидные структуры из сократительных белков актина и миозина, кот учавствуют: в обр цитоскелета и кл контактов; в обеспечении движение кл;
в локальных изменениях конфигурации ПАК; в транспорте в-в через ПАК; в сокращении мышечных волокон; в передвижении белков плазмолемы; в защите клетки от осмотического шока; в делении ЦП(цитокенез).
Располагаются под плазмолемой в кортикальном слое пучками или слоями. *Основу тонкой МФ составляет белок актин, кот имеет участки связывания с миозином и с белками тропомиозинового комплекса. К актинсвязывающим белками относят дистрофин, нарушение кот вызывает нарушение сократительной функции мышечных кл-миодистрофия Дюшена. Выделяют 3 формы актина: а-актин в кл скелетных мышц миокарда, в и у-актинв немыш кл-явл консервативными белками, кот способны к полимеризации(на одном конце фибрилла достраивается) и деполимеризации((разбирается на другом конце). G-актин в присутствии ионов Mg и АТФ собирается в полимерные формы-протофибриллы, кот обр фибрилярный F-актин. Между актиновыми спиралями расположены мол тропомиозина, обеспечивающие взаимод актина с миозином. *Толстые миозиновые МФ обр высокомол двигательным белком-миозином: мышечный(миозин II) и немышечный(миозин I). Миозин относится к белкам класса молекулярных моторов, способных перемещаться, преобразуя хим энергию гидролиза АТФ в механическую работу. Молекула миозина имеет головку(АТФ-азный центр(к головке присоед и гидролиз АТФ=>изм конформации головки и движение мол миозина по F-актину), актинсвязывающий центр, посредсвом кот головка миозина может прикрепляться к молекуле актина образуя хим. связи) и хвост. Миозин 2 сост из 6 полипептидов с 2-мя тяжелыми Н-цепями, 2 легкие L-цепи и 2 регуляторные цепи. Взаимод актина и миозина явл основой всех типов мышечного сокращения при увеличении конц ионов Са в ЦП, регулируемой спец кальциевых насосов.
Разновидность миозина 2 явл миозин 3, кот не способен формировать филаменты. Он обеспечивает движение вдоль актиновых МФ
Миозин 1 имеет одну глобулярную головку и более короткий стержень.
МФ обр в ООС два вида структур: сеть МФ и нити натяжения. Сеть МФ расположена на всём протяжении ПЛ, а нити натяжения представляют пучки МФ, кот предотвращают разрыв П под действием осмотического шока.
Промежуточные филаменты(ПФ) или скелетные фибриллы-тонкие нитевидные неветвящиеся структуры, часто располагаются пучками, выполняют опорную функцию(обр каркасили цитоскелет) и учавствуют в обр клеточных контактов.
ПФ состоят из специфических структурных белков. В эпительальных кл обр кератином и наз тонофибриллами, в нерв нкл ПФ-тремя разными белками(NF-1,NF-2,NF-3)и наз нейрофибрилами, а в мышечной ткани ПФ образованы белками-десминами. Белки ПФ представляют собой гомотетрамеры, т.е. состоят их 4идентичных полипептидов.
ПФ обр путём полимеризации соответсвующих тетрамерных белков ПФ. Устойчивость к действию хим факторов, например к алкоголю(у алкоголиков в гепатоцитах МФ и МТ заменяются на скелетные фибрилы). ПФ явл компонентом ЦП и ядра, участвуют в формировании цитоскелета кл.
Микротрубочки(МТ)-полые белковые структуры, состоящие из 3 видов полимеризующихся беков-тубулинов(а,В,у). Сборка МТ начинается в ЦП в центрах организации МТ. В присутствии ионов Mg, АТФ и у-тубулина в качестве затравки,а- и В-тубулины обр стабильные гетеродимеры. В результате а-тубулинодного димера взаимодействует с В-тубулином другог димера и наоборот. Затем 13 тубулиновых протофиламентов приводят к образованию1 МТ.Удлинение МТ осущ на 2 концах. Быстро растущий «плюс-конец», медленно «минус-конец».
На процессы полимеризации и деполимеризации влияют: повышение давления и понижение температуры(разрушение), алкоголь, растительный алколоид колхицин,
МТ могут содержать нетубулиновые белки, кот выполняют структурную или регуляторную функции.
Кроме транспорта мембранных белков МТ учавствуют в ЦП(транспорте в-в в мембранной упаковке в клеткуи из кл), в обр опоры к или цитоскелета, определяют их форму и строят нити веретена деления.
2..Трипаносомы(протозоа.простейшие)



3. Генетика человека. Близнецовый метод. Цель и задачи.
Среди методов генетического анализа большое значение имеет близнецовый метод, позволяющий отдифференцировать роль условий среды и генотипа в развитии различных признаков, предрасположения к заболеваниям и др.
Суть метода в сравнении разных групп близнецов, исходя из сходства и различия их генотипов и среды; в которой они росли. При этом сопоставляют:
монозиготных близнецов с дизиготными;
монозиготных близнецов между собой;
результаты анализа близнецовой выборки в общей популяции.
Близнецы могут быть однояйцевыми (монозиготными, идентичными) или разнояйцевыми (дизиготными, неидентичными). Однояйцевые близнецы возникают на самых ранних стадиях дробления зиготы, когда два или четыре бластомера при обособлении сохраняют способность развиться в полноценный организм. Зигота делится митозом, поэтому генотипы однояйцевых близнецов идентичны. Однояйцевые близнецы всегда одного пола.
Разнояйцевые близнецы возникают при оплодотворении двух или нескольких одновременно созревших клеток. Они имеют около 50% общих генов, т.е. подобны обычным братьям и сестрам, рожденным в разное время, и могут быть однополыми и разнополыми. Частота рождения близнецов невелика и составляет около 1% (1/3 однояйцевые, 2/3 разнояйцевые).
Большинство близнецов - двойни. Многоплодные беременности бывают не только двойнями, но и тройнями, четвернями и более.
Исследование с использованием близнецового метода состоит из трех этапов:
1. Составление выборки. Для этого в популяциях или отбирают всех
близнецов, а затем тех, кто имеет анализируемые признаки, или из всего населения
выделяют лиц с данными признаками, а потом среди них - близнецов.
2. Установление зиготности. В основе диагностики зиготности лежит изучение
сходства (конкордантности) и различия (дискордантности) партнеров близнецовой
пары по совокупности таких признаков, которые изменяются под
влиянием среды Сравнивают детей по совокупности внешних признаков
(пигментация кожи, волос, форма носа, рук, губ и др.)
3. Монозиготные близнецы конкордантны по всей совокупности признаков, а дизиготные по некоторым признакам могут быть дискордантны.
Другой метод установления зиготности - иммуногенетический. Близнецов сравнивают по эритроцитарным антигенам системы АВ0, Rh и др., и составу белков сыворотки.
Эти менделирующие признаки не изменяются в течение жизни и не зависят от внешних факторов. При отсутствии ошибок определения даже единственное различие может свидетельствовать о дизиготности близнецов.
Используют также метод дерматоглифики (исследование кожных узоров пальцев рук и ладоней) (3. Восстановление пар и групп близнецов по рассматриваемым признакам.
Методы сравнения близнецовых выборок по качественным (дискретным) признакам (ахондроплазия, альбинизм и др.) и количественным (рост, масса тела, артериальное давление, продолжительность жизни и др.) различны.
Степень конкордантности по качественным признакам у монозиготных близнецов очень высока и стремится к 100%, а у дизиготных меньше - около 70%.
При изучении роли наследственности и среды в формировании количественных признаков степень различия близнецов определяется у дизигот -наследственностью и средой, у монозигот - только средой.
При идентичном генотипе сходная реакция на внешний фактор (туберкулезная инфекция) наступает чаще, чем при разных генотипах, что доказывает важную роль генетических факторов. Исследования показали, что высокая конкордантность способствует не только возникновению болезни, но и ее клиническому проявлению
Из количественных признаков рост меньше зависит от влияния факторов внешней среды, чем масса. Различие роста между монозиготными близнецами составляет 1,7 см, между дизиготными - 4,4 см.
Монозиготные близнецы значительно чаще болеют формами туберкулеза, одинаковыми по течению и исходу. Монозиготные близнецы обычно проявляют способности к одному и тому же виду деятельности, а дизиготные - к различным.
Близнецовый метод позволил доказать основной закон генетики развития: индивидуальные свойства каждого организма формируются в онтогенезе под контролем генотипа и среды.
Действие факторов среды на развитие признаков после рождения можно проследить в том случае, если сразу после рождения их воспитывать отдельно.
Например, один живет в городе, а другой - в сельской местности. Если близнецы монозиготны, то можно определить влияние факторов среды в формировании признаков, составить представление о норме реакции данного гена. При сравнении моно- и дизиготных близнецов в одной и той же среде можно сделать вывод о роли генов в развитии признаков.
Близнецовый метод позволил применить метод контроля по партнеру. В этом методе используют только монозиготных близнецов. Зная, что генотипы их одинаковы, можно точно оценить действие внешнего фактора на одного из них, при условии если другой не подвергается этому воздействию и служит контролем. Если монозиготные близнецы конкордантны по болезни, и один из них получает новый лечебный препарат, а другой служит "контролем", то это позволяет получить объективные сведения об эффективности препарата. Метод контроля по партнеру применяют в клинической генетике и фармакологии.
Сравнение однояйцевых и разнояйцевых близнецов имеет значение для изучения влияния генотипа и среды в развитии определенного фенотипа. Для качественных признаков используется конкордантность(наличие признака у обоих близнецов) и дисконкордантность.
Формула Хольцингера:
Н=(Коб-Крб)/(1-Крб)=(Дрб-Доб)/Дрб
Н-коэф.наследуемости
Н=0.7 и более, ведущую роль в развитии данного признака играет генотип
Н<0.7 ведущую роль играют факторы внешней среды и этот признак имеет широкую норму реакции.
4. Естественный отбор и его формы, дизруптивный, движущий, стабилизирующий.
Движущий отбор — форма естественного отбора, которая действует при направленном изменении условий внешней среды. В результате в популяции из поколения к поколению происходит сдвиг средней величины признака в определённом направлении. При этом давление движущего отбора должно отвечать приспособительным возможностям популяции и скорости мутационных изменений
Стабилизирующий отбор — форма естественного отбора, при которой его действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака. Например: серповидно-клеточная анемия
Дизруптивный (разрывающий) отбор — форма естественного отбора, при которой условия благоприятствуют двум или нескольким крайним вариантам (направлениям) изменчивости, но не благоприятствуют промежуточному, среднему состоянию признака. В результате может появиться несколько новых форм из одной исходной.