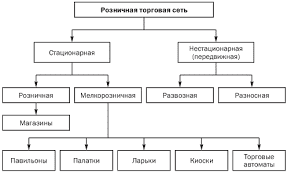
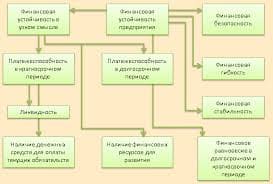
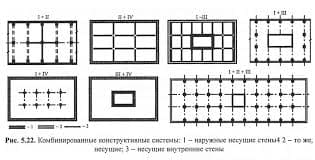
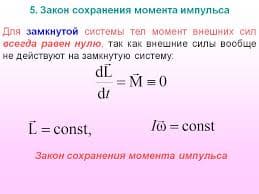2020-04-20
2020-04-20 237
237
Первые попытки этого синтеза начались с конца 30‑х и продолжались в течение 40‑х годов; они были плодом коллективных усилий биологов разных специальностей.
Дж. Симпсон[136] писал, что среди многих соавторов синтетической теории эволюции необходимо отметить в Англии Фишера, Холдейна, Хаксли, Дарлингтона, Уоддингтона и Форда, в США‑Райта, Мёллера, Добжанского, Майра, Дайса и Стеббинса, в Советском Союзе – Четверикова, Дубинина, Тимофеева‑Ресовского, в ФРГ – Ренша, во Франции – Тейсье, в Италии – Бузати‑Траверзо. Ф.Г. Добжанский и Э. Бозигер[137] привели 15 имен, в числе которых, кроме части упомянутых Симпсоном, названы Грант, Симпсон, Шмальгаузен и Уайт. Ж. Тейсье[138] упомянул примерно 20 ученых.
Перечисление исследователей, осуществивших синтез теории естественного отбора с генетикой и экологией, справедливо было бы продолжить и помимо уже названных биологов отметить также и заслуги таких ученых, как Н.И. Вавилов, Г.Д. Карпеченко, Дж. Клаузен, И. Лернер Ф. Шеппард, К. Мазер, К. Сакаи, М.А. Розанова, М.М. Камшилов, Г.Ф. Гаузе, В.Н. Сукачев, Ч. Элтон, Дж. Лэк, Т. Парк, Д.Н. Кашкаров, Е.Н. Синская, С.А. Северцов, Г. Турессон. Таким образом, в создании основ современного дарвинизма участвовало, видимо, не менее 40–50 ученых из восьми стран.
К наиболее важным работам, положившим начало формированию синтетической теории эволюции, следует отнести не только труд Добжанского «Генетика и происхождение видов» (1937), но и серию обобщающих работ И.И. Шмальгаузена – «Организм как целое в индивидуальном и историческом развитии» (1938), «Пути и закономерности эволюционного процесса» (1939) и «Факторы эволюции» (1946). Существенное значение в создании синтетической теории эволюции имеют книги Н.В. Тимофеева‑Ресовского «Генетика и эволюция» (1939), С. Дарлингтона «Эволюция генетических систем» (1939), Дж. Хаксли «Эволюция. Современный синтез» (1942), Э. Майра «Систематика и происхождение видов» (1942), Дж. Симпсона «Темпы и формы эволюции» (1944), работы Н.П. Дубинина с сотрудниками (1936–1948).
В современной синтетической теории главным фактором эволюции признается естественный отбор, интегрирующий и регулирующий действие всех остальных факторов (онтогенетической изменчивости, мутагенеза, гибридизации, миграции, изоляции, пульсации численности и др.). Понятно поэтому, что естественный отбор служит объектом всесторонних исследований.
Важная особенность синтетической теории эволюции состоит в замене типологического (организмоцентрического) подхода к пониманию субстрата эволюции качественно новым популяционным подходом, основывающимся на признании элементарной единицей эволюции местной (локальной) популяции. Основу синтетической теории составляют не данные эволюции крупных систематических групп, интерпретация которых неизбежно включает много гипотетического, а факты, получаемые в исследованиях микроэволюции. Эти факты добываются Путем непосредственного экспериментального исследования элементарных процессов, протекающих в популяциях. Это не означает, что новый синтез не касается проблем макроэволюции. Образование крупных таксонов, органообразование, направленность эволюции и другие вопросы макроэволюции продолжают оставаться в поле ее зрения, но фундаментом для истолкования этих данных служат точно установленные факты, объясняющие причины микроэволюции.
Для синтетической теории эволюции характерно использование комплексных методов исследования: экспериментальных в сочетании с моделированием различных моментов эволюционного процесса, сравнительно‑географических и экологических опытов в природной обстановке и в питомниках, полевых наблюдений, данных, добытых сравнительно‑морфологическими методами, а также широкое применение математики. Использование этих методов позволило подтвердить правильность главных положений дарвинизма о ведущей роли естественного отбора, о том, что ни один из отдельно взятых факторов эволюции не объясняет ее причин, являющихся результатом сложных взаимодействий этих факторов. Новая теория разработала эти положения применительно к различным объектам, начиная от бактерий и простейших и кончая высшими растениями и животными.
После строгой экспериментальной проверки были опровергнуты некоторые концепции (наследование приобретенных признаков, автогенез и эктогенез, преадаптационизм и др.), а также преодолена недооценка роли модификационной изменчивости и фенотипа в эволюции. Статистическое представление о движущих силах эволюции было приведено в соответствие с учением о популяции как субстрате эволюции. Новая теория включила в себя все крупные достижения эволюционизма последарвиновского времени. Многие факты, оказавшиеся трудными для объяснения с позиций классического дарвинизма и использовавшиеся для его критики (например, вредность мутаций, явление преадаптации, неадаптивный характер некоторых эволюционных изменений) были подчинены теории естественного отбора и получили рациональное объяснение.
Ранее казалось незыблемым положение, что наследственная изменчивость является причиной, а отбор – лишь следствием, действующим на ее основе. В работах 1936–1946 гг. Н.П. Дубинин, Н.И. Шапиро, А. Стертевант, М.В. Игнатьев, Р.Л. Берг и другие показали, что сам мутационный процесс является адаптивным признаком вида; этот признак преобразуется отбором, который изменяет мутабильность, поддерживая ее на оптимальном уровне. Стало ясно, что мутационный процесс является не только исходным материалом эволюции, но и ее результатом.
Концепция «мобилизационного резерва» внутривидовой наследственной изменчивости (И.И. Шмальгаузен, С.М. Гершензон и другие) позволила снять одно из затруднений, стоявших перед теорией естественного отбора. Шмальгаузен (1946) показал, что «процесс вскрытия уже существующих резервов имеет гораздо большее значение, чем самая интенсивная мутабильность, так как мутации, ранее накопленные в скрытом виде, проявляются сразу в значительной концентрации, обеспечивая тем самым наиболее высокую эффективность естественного отбора»[139].
Было показано, что рецессивность, доминантность и сверхдоминантность – это итоги деятельности отбора, направленного на устранение вредящего действия мутаций, на образование мобилизационного резерва, делающего вид эволюционнопластичным, на формирование адаптивного фенотипа, на его упрочение и защиту от дезорганизующего действия новых мутаций (Р. Гольдшмидт, М.М. Камшилов, Д.М. Шифрина и другие).
Был выяснен механизм, поддерживающий невысокую численность популяций бабочек, подражающих несъедобным насекомым; оказалось, что их малочисленность является необходимым условием выживания, так как при большей численности исчезает «эффект примеси», и птицы начали бы отличать имитаторов от моделей (Е. Форд, Г. Кэттлуэлл, Р. Шеппард и другие). Численность подобных видов регулируется летальными генами, концентрация которых бывает весьма высокой (гибнет до 60–80 % зародышей в каждом поколении). Но именно эта постоянная высокая смертность является важной адаптацией вида. У многих животных и растений были обнаружены гены, летальные или снижающие жизнеспособность в гомозиготном состоянии, а в гетерозиготном – повышающие жизнеспособность, иммунитет, экологическую пластичность и другие важнейшие показатели. Оказалось, что отбор часто сохраняет генетические системы, способные тем или иным способом поддерживать состояние адаптивной гетерозиготности, и действует против гомозигот.
При изучении полиморфизма удалось выяснить роль отбора, поддерживающего постоянство численных отношений между различными формами в популяциях (Дубинин, Добжанский, Шеппард и другие). Но при известных условиях устойчивый (сбалансированный) полиморфизм может смениться переходным, при котором отбор будет способствовать преобладанию одной (или нескольких) формы над другими, вплоть до полной качественной перестройки состава популяции (Форд и другие).
Состояние эволюционной теории в конце первой половины XX в. получило отражение в содержании работы симпозиума, посвященного экологическим и генетическим факторам видообразования и эволюции (1949, Италия). На этом симпозиуме обсуждались многие вопросы, но «в центре внимания находилась проблема естественного отбора»[140]. В ряде докладов было показано, что отбор имеет дело не с отдельными генами или признаками, а элиминирует только целые фенотипы недостаточной адаптивной ценности, развивающиеся на базе целых генотипов. Было отмечено, что у высокоадаптивных форм могут развиваться отдельные нейтральные или даже слабо вредные признаки. Подробно обсуждалось положение о том, что генотип вида является целостной системой, постоянно интегрируемой естественным отбором, и критиковалось представление о генотипе как простой сумме независимых генов. Высказывалась мысль, что при образовании морфологически неразличимых, но репродуктивно изолированных популяций отбор охраняет морфологическую структуру как адаптивно особенно ценную, и дивергенция популяций идет только по физиологическим признакам. Участники симпозиума резюмировали, что естественный отбор всегда оказывается решающей причиной видообразования и общим направляющим фактором эволюции.
Важнейшей тенденцией развития эволюционной теории в 50‑60‑е годы было расширение и углубление синтеза биологических знаний на основе изучения биологии популяций и закономерностей естественного отбора.
Значительно усилилось взаимодействие между эволюционной теорией и такими отраслями биологии, как генетика популяций, популяционная экология, этология, биогеография и микросистематика.