 2014-02-05
2014-02-05 911
911Таблица 3.9 – Модель хода роста древостоя еловых культур № 4
Для предотвращения падения производительности и будущих запасов древесины необходимы рубки, снижающие густотуперед пикомсомкнутости крон, а не на его максимуме, с удалением нередко более половины живых деревьев в возрасте до 20-25 лет.
Одноярусные древостои всех пород, составов, различной начальной густоты и неравномерности расположения деревьев имеют своей целью общее стремление всех деревьев яруса к достижению предельных линейных, площадных и объемных показателей в пределах сообщества. В результате почти любой древостой однажды в жизни становится сомкнутым и достигает индивидуального предельного состояния по этим показателям.
Но если эта оценка меняется за 40 лет на 2-3 градации бонитета или до 60%, то следует ли ее применять для оценки производительности условий местопроизрастания, которые на самом деле не менялись?
Ответы на эти неудобные вопросы не получили иного объяснения кроме рабочей гипотезы Г.С.Разина о том, что изменения в ходе роста древостоев вызваны главным образом различиями в их начальной густоте. С этого предположения, собственно, и начались исследования в направлении, которые закончились открытием нижеследующих закономерностей по сомкнутости древостоев.
|
|
|
Рост, развитие и возрастная динамика древостоев подчиняются некоторым всеобщим биологическим законам. В дендроценозах эти законы имеют специфические особенности.
2. Достижение предельных показателей происходит в зависимости от густоты растений и может быть сформулировано в виде «Закона динамики суммы горизонтальных проекций крон деревьев в простых древостоях», который показан в виде нескольких линий зависимости на рисунке 4.1а и состоит в следующем:
- чем больше начальная густота, тем раньше и при меньшей средней высоте древостои становятся сомкнутыми с предельной суммой площадей горизонтальных проекций крон деревьев и тем раньше и быстрее они снижают ее (размыкаются);
- чем меньше начальная густота, тем позже и при больших средних высотах древостои достигают предел сомкнутости и предел суммы площадей горизонтальных проекций крон деревьев, тем медленнее они снижают их и дольше остаются сомкнутыми и устойчивыми.
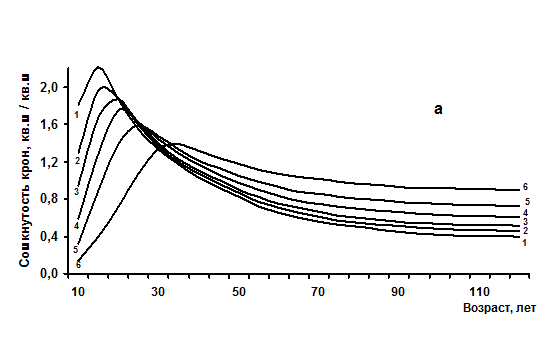
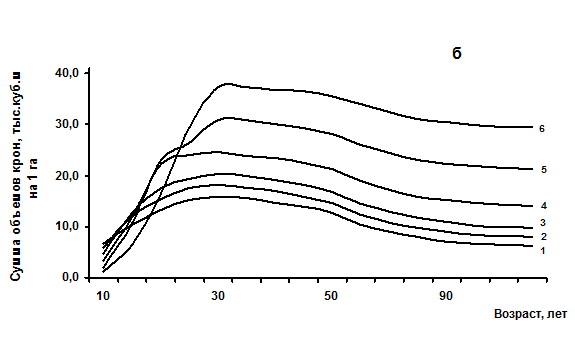
Рис. 4.1 – Возрастная динамика сомкнутости крон (а) и объемов крон (б) одноярусных еловых древостоев с различной начальной густотой (в возрасте 10 лет), тыс. шт/га: 1 - 67; 2 – 22; 3 – 11,5; 4 – 7; 5 – 4,5; 6 – 1,8 тыс. шт/га.
Открытый для площадных показателей закон проявляет себя аналогичным образом в показателях объема. С некоторым запаздыванием по сравнению с сомкнутостью (на 10 – 25 лет) древостои точно также достигают предельного состояния по сумме объемов крон и имеют индивидуальные линии своего развития по этому показателю в зависимости от начальной густоты (рис. 4.1б).
|
|
|
Интерпретация в биологическом плане обнаруженных законов выглядит следующим образом. Возникшие одноярусные сообщества древесных растений обладают сильнейшим изначальным свойством (способностью) у каждого члена сообщества, которое можно назвать целеполаганием или «целью» – стремлением к достижению индивидуальных пределов по протяженности (высоте, длине, толщине), а также по площади и по объему.
Важно отметить, что при любых высотах и возрастах в ельниках бывают состояния с предельной сомкнутостью. При средней высоте 1 м сумма площадей крон деревьев может быть в 2,6 раза больше площади поверхности, на которой они расположены (предельная сомкнутость 2,60) и кроны деревьев в этом случае заходят в кроны второго и даже третьего дерева по соседству. При высоте 20 м сомкнутость имеет предел 1,24. Достижение таких предельных значений воспринимается наблюдателем как древостой с почти полным отсутствием растительности в напочвенном покрове; тип леса определить затруднительно и условно он может быть назван «мертвопокровным».


В интегрированном виде описанные выше законы могут быть объединены в общий закон развития одноярусных древостоев, который утверждает: чем больше начальная густота древостоев, тем меньше по размерам и объему в них деревья; такие древостои хуже растут и с возрастом оказываются менее полными и низкопроизводительными по сравнению с древостоями с меньшей начальной густотой. Густые древостои плохо противостоят факторам стресса и быстрее распадаются по сравнению с менее густыми по своим внутренним причинам (Разин, Рогозин, 2010).
Следствием данного закона является перегруппировка древостоев по мере их развития по всем таксационным показателям: сомкнутости, полноте, сумме объемов крон деревьев, запасам древесины, общей производительности и классам бонитета. Рассмотрим производительность насаждений в моделях хода роста елового древостоя.
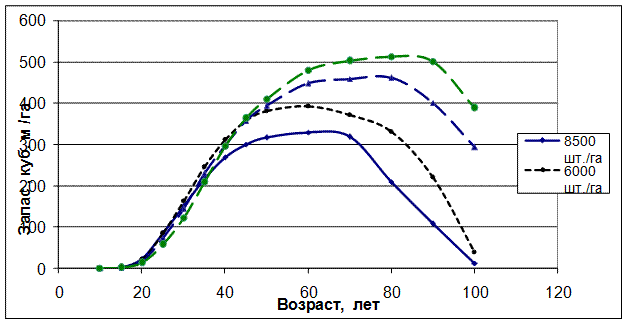 РРис. 3.3 – Запасы древесины в моделях хода роста культур ели
РРис. 3.3 – Запасы древесины в моделях хода роста культур ели
различной начальной (в 10 лет) густоты.
В соответствии с обнаруженным законом можно утверждать, что начальная густота древостоев и возникающая конкуренция между растениями в решающей степени определяют всю последующую жизнь насаждения и чем больше густота, тем сильнее меняется рост и развитие как отдельных деревьев, так и древостоев в целом.
Густота оставшихся деревьев должна быть равномерной и оптимальной (см. формулу Разина)

Изреживание культур ели в возрасте 21 год с оставлением оптимального количества деревьев на единице площади, равномерно размещенных на ней.



| Таблица 3.14 – Пермский край, южная и средняя подзоны тайги |
| ТУМ С3,В3. Типы леса: Ечерн., Етрав., Езм., Ек-зм. (в 40-70 лет мертвопокровные) |
| В 25 лет – начало рубок ухода слабой и умеренной интенсивности |
| Весь растущий древостой | Отпад и промежуточное пользование | Общая | ||||||||||||||||
| производительность | ||||||||||||||||||
| А, лет | Дср, см | Нср, м | Дср/Нср, см/м | Nтек, шт/га | ∑g, м2/га | HF, м3/м2 | М, м3/га | ΔМср, м3/га | ΔМтек, м3/га | Бони-тет | Пол-нота | Nотп, шт/га | Vср.отп, м3/шт. | Мотп, м3/га | Побщ, м3/га | ΔПср, м3/га | ΔПтек, м3/га | |
| 0,8 | 0,2 | 0,02 | 0,02 | V | 0,2 | 0,02 | 0,02 | |||||||||||
| 1,2 | 1,6 | 1,31 | 0,4 | 2,19 | 0,9 | 0,06 | 0,14 | lV,9 | 0,11 | 0,9 | 0,06 | 0,14 | ||||||
| 4,5 | 3,5 | 1,29 | 5,49 | 2,33 | 12,8 | 0,64 | 2,4 | lV,0 | 0,47 | 0,0001 | 12,8 | 0,64 | 2,4 | |||||
| 7,5 | 6,6 | 1,14 | 3,81 | 2,3 | 8,9 | lll,2 | 0,63 | 0,0015 | 0,1 | 57,1 | 2,3 | 8,9 | ||||||
| 12,5 | 10,4 | 1,20 | 20,7 | 5,26 | 3,6 | 10,4 | ll,0 | 0,56 | 0,0073 | 12,5 | 4,1 | 13,0 | ||||||
| 16,7 | 13,8 | 1,21 | 27,5 | 6,47 | 5,1 | 13,8 | l,2 | 0,59 | 0,0304 | 13,2 | 5,8 | 16,4 | ||||||
| 19,2 | 15,9 | 1,21 | 36,3 | 7,36 | 6,7 | 17,8 | l,0 | 0,71 | 0,0727 | 7,3 | 17,8 | |||||||
| 23,0 | 18,5 | 1,24 | 35,9 | 8,44 | 6,7 | 7,2 | lа,7 | 0,63 | 0,117 | 8,3 | 16,4 | |||||||
| 24,3 | 1,22 | 40,1 | 9,35 | 7,5 | 7,2 | lа,7 | 0,68 | 0,206 | 8,9 | 14,4 | ||||||||
| 28,5 | 22,4 | 1,27 | 41,1 | 10,4 | 7,1 | 5,2 | lа,8 | 0,65 | 0,290 | 9,4 | 11,6 | |||||||
| 30,5 | 24,2 | 1,26 | 47,1 | 11,3 | 7,6 | 10,5 | lа,9 | 0,72 | 0,494 | 9,5 | 10,6 | |||||||
| 33,5 | 25,7 | 1,30 | 12,1 | 6,8 | 1,2 | l,0 | 0,68 | 0,660 | 9,6 | 10,4 | ||||||||
| 35,7 | 26,7 | 1,34 | 51,1 | 12,5 | 7,1 | 9,5 | l,2 | 0,76 | 0,961 | 9,6 | 9,5 | |||||||
| 37,4 | 27,5 | 1,36 | 13,0 | 7,3 | 8,9 | l,3 | 0,83 | 1,228 | 9,5 | 8,9 | ||||||||
| 39,0 | 27,8 | 1,40 | 60,9 | 13,1 | 7,2 | 7,0 | l,5 | 0,9 | 1,510 | 9,3 | 7,0 |
|
|
|








