 2014-02-09
2014-02-09 1179
1179Молекулярно-генетический уровень.
Элементарной единицей на молекулярно-генетическом уровне служит ген - фрагмент молекулы нуклеиновой кислоты, в котором записан определенный в качественном и количественном отношении объем биологической (генетической) информации. Элементарное явление заключается, прежде всего, в процессе конвариантной редупликации, или самовоспроизведении, с возможностью некоторых изменений в содержании закодированной в гене информации. Путем редупликации ДНК происходит копирование заключенной в генах биологической информации, что обеспечивает преемственность и сохранность (консерватизм) свойств организмов в ряду поколений.
В силу ограниченной стабильности молекул или ошибок синтеза в ДНК (время от времени, но неизбежно) случаются нарушения, которые изменяют информацию генов. В последующей редупликации ДНК эти изменения воспроизводятся в молекулах – копиях и наследуются организмами дочернего поколения. Указанные изменения возникают и тиражируются закономерно, что и делает редупликацию ДНК конвариантной, т.е. происходящей иногда с некоторыми изменениями. Такие изменения в генетике получили название генных (или истинных) мутаций. Конвариантность редупликации, таким образом, служит основой мутационной изменчивости.
Биологическая информация, заключающаяся в молекулах ДНК, не участвует непосредственно в процессах жизнедеятельности. Она переходит в действующую форму, будучи перенесена в молекулы белков. Отмеченный перенос осуществляется благодаря механизму матричного синтеза, в котором исходная ДНК служит, как и в случае с редупликацией, матрицей, но для образования не дочерней молекулы ДНК, а матричной РНК, контролирующей биосинтез белков. Отмеченное дает основание причислить матричный синтез информационных макромолекул к элементарному явлению на молекулярно-генетическом уровне организации жизни.
Нуклеиновые (от лат. nucleus — ядро) кислоты — сложные органические соединения. Они состоят из углерода, водорода, кислорода, азота и фосфора.
Существуют два типа нуклеиновых кислот — ДНК и РНК. Они могут находиться как в ядре, так и в цитоплазме и ее органоидах.
ДНК — дезоксирибонуклеиновая кислота. Это биологический полимер, состоящий из двух полинуклеотидных цепей, соединенных друг с другом. Мономеры — нуклеотиды, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения. ДНК состоит из четырех азотистых оснований: производных пуринов — аденина (А) и гуанина (Г) и производных пиримидинов — цитозина (Ц) и тимина (Т), пятиатомного сахара пентозы — дезоксирибозы, а также остатка фосфорной кислоты (рис.).
В каждой цепи нуклеотиды соединяются между собой ковалентными связями: дезоксирибоза одного нуклеотида соединяется с остатком фосфорной кислоты последующего нуклеотида. Две цепи объединяются в единую молекулу водородными связями, возникающими между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи. Пространственная конфигурация азотистых оснований различна и количество таких связей между разными азотистыми основаниями неодинаково. Вследствие этого они могут соединяться только попарно: азотистое основание аденин (А) одной цепочки пол и нуклеотида всегда связано двумя водородными связями с тимином (Т) другой цепи, а гуанин (Г) — тремя водородными связями с азотистым основанием цитозином (Ц) противоположной полинуклеотидной цепочки. Такая способность к избирательному соединению нуклеотидов, в результате которого формируются пары А—Т и Г—Ц, называется комплементарностью (рис. 15). Если известна последовательность оснований в одной цепи (например, Т —Ц—А—Т—Г), то благодаря принципу комплементарности (дополнительности) станет, известна и противоположная последовательность оснований (А-Г-Т-А-Ц).
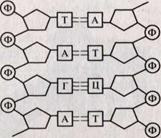
Рис. Участок молекулы ДНК. Комплементарное соединение нуклеотидов разных цепей.
Цепи нуклеотидов образуют правозакрученные объемные спирали по 10 оснований в каждом витке. Последовательность соединения нуклеотидов одной цепи противоположна таковой в другой, т.е. цепи, составляющие одну молекулу ДНК, разнонаправленны, или антипараллельны: последовательность межнуклеотидных связей в двух цепях направлена в противоположные стороны: 5' —3' и 3' —5'. Сахаро-фосфатные группировки нуклеотидов находятся снаружи, а азотистые основания — внутри. Цепи, закрученные друг относительно друга, а также вокруг общей оси, образуют двойную спираль. Такая структура молекулы поддерживается в основном водородными связями (рис.).
Вторичную структуру ДНК впервые установили американский биолог Дж.Уотсон и английский физик Ф. Крик.


Рис. Схема строения двойной спирали ДНК: А — плоскостная модель, жирной чертой обозначен сахаро-фосфатный остов; Б — объемная модель
При соединении ДНК с определенными белками (гистонами) степень спирализации молекулы повышается - возникает суперспиральДНК, толщина которой существенно возрастает, а длина сокращается (рис.). Единицей компактизации молекулы ДНК служит нуклеосома, основу которой составляют 8 молекул гистонов, по 2 каждого типа (Н2А, Н2В, НЗ и Н4). Поверхности этих белковых молекул несут положительные заряды и образуют остов, вокруг которого может закручиваться отрицательно заряженная молекула ДНК. В каждую нуклеосому входит от 146 до 200 пар нуклеотидов. Гистон пятого типа - Н1 - соединяется с участками ДНК, связывающими одну нуклеосому с другой.. Такая ДНК носит название линейной или соединительной — линкерной. Нуклео
сомы расположены вдоль ДНК на определенном расстоянии,неодинаковом в зависимости от типа клеток — от 20 до 50 нм. Так образуется структура, похожая на бусы, где каждая бусина — нуклеосома.
Рис.. Схема образования суперспирали ДНК.
30 нм








