 2014-02-12
2014-02-12 7216
7216Твердое тело может быть как кристаллическим, так и аморфным. В первом случае имеется дальний порядок в расположении частиц на расстояниях, много превышающих межмолекулярные расстояния (кристаллическая решетка). Во втором – нет дальнего порядка в расположении атомов и молекул.
Различие между аморфным телом и жидкостью состоит не в наличии или отсутствии дальнего порядка, а в характере движения частиц. Молекулы жидкости и твердого тела совершают колебательные (иногда вращательные) движения около положения равновесия. Через некоторое среднее время («время оседлой жизни») происходит перескок молекул в другое положение равновесия. Различие заключается в том, что «время оседлой жизни» в жидкости намного меньше, чем в твердом состоянии.
Липидные двухслойные мембраны при физиологических условиях – жидкие, «время оседлой жизни» фосфолипидной молекулы в мембране составляет 10−7- 10−8 с.
Молекулы в мембране расположены не беспорядочно, в их расположении наблюдается дальний порядок. Фосфолипидные молекулы находятся в двойном слое, а их гидрофобные хвосты примерно параллельны друг другу. Есть порядок и в ориентации полярных гидрофильных голов.
Физиологическое состояние, при котором есть дальний порядок во взаимной ориентации и расположении молекул, но агрегатное состояние жидкое, называется жидкокристаллическим состоянием. Жидкие кристаллы могут образовываться не во всех веществах, а в веществах из «длинных молекул» (поперечные размеры которых меньше продольных). Могут существовать различные жидкокристаллические структуры (рис.4.4, б, в, г): нематическая (нитевидная), когда длинные молекулы ориентированы параллельно друг другу; смектическая – молекулы параллельны друг другу и располагаются слоями; холестическая – молекулы располагаются параллельно друг другу в одной плоскости, но в разных плоскостях ориентации молекул разные.

Рис.4.4. Расположение молекул в аморфном (а), и жидкокристаллическом состоянии (б, в, г).
Двуслойная липидная фаза мембран соответствует смектическому жидкокристаллическому состоянию.
Жидкокристаллические структуры очень чувствительны к изменению температуры, давления, химического состава, электрического поля. Например, при понижении температуры в фосфолипидной мембране происходит переход из жидкокристаллического в гель-состояние (рис.4.5). В гель-состоянии молекулы расположены еще более упорядоченно, чем в жидкокристаллическом. Все углеводородные гидрофобные хвосты фосфолипидных молекул в гель-фазе полностью вытянуты строго параллельно друг другу (имеют полностью транс-конформацию). В жидком кристалле за счет теплового движения возможны транс-гош-переходы, хвосты молекул изгибаются, их параллельность в отдельных местах нарушается и особенно сильно в середине мембраны.
Толщина мембраны в гель-фазе больше, чем в жидком кристалле. Однако при переходе из твердого в жидкокристаллическое состояние объем несколько увеличивается, потому что возрастает площадь мембраны (в расчете на одну молекулу площадь возрастет с 0,48 до 0,58 нм2). Так как в твердокристаллическом состоянии упорядоченность в строении выше, чем в жидком кристалле, ему соответствует меньшая энтропия.
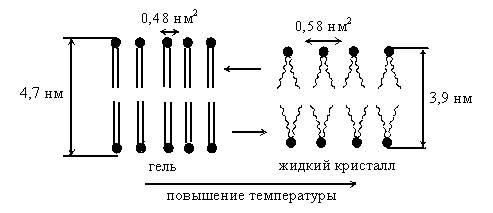
Рис.4.5.Изменение структуры мембраны при переходе из жидкокристаллического состояния в гель-состояние и обратно при изменении температуры.
Для нормального функционирования мембрана должна быть в жидкокристаллическом состоянии. Поэтому в живых системах при продолжительном понижении температуры окружающей среды наблюдается адаптационное изменение химического состава мембран в результате обмена веществ. Это обеспечивает понижение температуры фазового перехода. Температура фазового перехода понижается при увеличении числа ненасыщенных связей в составе жирных кислот в фосфолипидах. Температура фазового перехода гель – жидкий кристалл может изменяться от – 20 ºС (для мембран из ненасыщенных липидов) до + 60 ºС (для насыщенных липидов). Увеличение числа ненасыщенных липидов в мембране при понижении температуры среды обитания наблюдается у всех организмов.
Гипотеза петли (кинка). Плавление жирнокислотных цепей при фазовом переходе обусловлено вращательной изомерией. Наименьшей энергией обладает транс-, а наибольшей - цис-конформация. Гош-конформации [гош (+) и гош (−) - это поворот на ±120° относительно транс – конформации]. Синоним гош-конформации – скошенная конформация. Он сравнительно мало превышают по энергии транс-конформацию (на 2-3 кДж/моль), но эти состояния разделяет энергетический барьер высотой ~ 12-17 кДж/моль. Если углеводородные цепи, находящиеся полностью в транс-конформации представляют собой линейные структуры, то появление одиночной гош-конформации в цепи приводит к искривлению пространственной конфигурации цепи на угол ~ 120°. В плотно упакованных мембранных системах с полностью транс-конформацией углеводородных цепей это искривление порождает серьезные стереохимические затруднения, делающие невозможным появление одиночных гош-конформаций.
Уменьшение стереохимических затруднений при плавлении углеводородных цепей в мембранах достигается при синхронном появлении в цепи сразу двух гош-конформаций [гош (+) и гош (−)], разделенных одиночной С − С-связью в транс-конформации. Хотя энергия такого состояния вдвое выше одиночной гош-конформации, возникает ротационное состояние цепи. Оно не вызывает сильного расширения решетки, так как при последовательном повороте цепи на +120 и −120° пространственная конфигурация цепи в целом сохраняется прямолинейной (рис.4.6, А). Участок цепи, находящийся в гош (+)-транс-гош (−)-конформации, формирует уступ или петлю в углеводородной цепи, которую часто называют кинком (от английского kink —петля).
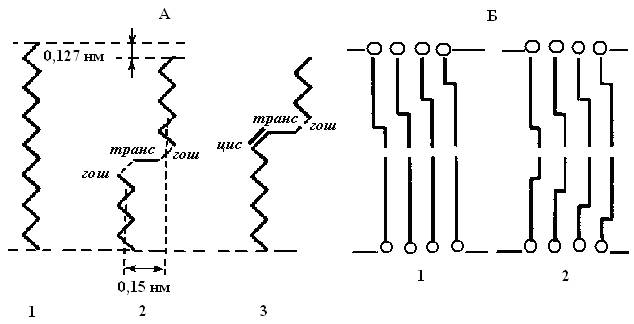
Рис.4.6. А: 1 - углеводородные цепи полностью в транс-конформации, 2 - в гош-транс-гош-конформации, 3 – в цис-транс-гош-конформации; Б: кинк-блоки в углеводородных цепях мембран: 1 - в одном монослое мембраны; 2 – в двух монослоях липидного бислоя.
Образование кинка сопровождается уменьшением эффективной длины цепи на ~ 0,127 нм. При этом часть цепи отодвигается на ~ 0,15 нм, образуя свободный объем, а занимаемый молекулой липида общий объем увеличивается на 0,025-0,050 нм3. Хотя появления одного кинка в углеводородной цепи недостаточно для ее плавления, однако одиночные кинки облегчают возникновение кинков в соседних углеводородных цепях, формируя чередующиеся кинк-блоки (рис. 4.6, Б). Такие блоки могут возникать либо в одном монослое мембраны, либо в двух противоположно расположенных углеводородных слоях (в бислое). При увеличении числа кинков в углеводородных цепях неупорядоченность углеводородной зоны мембран резко нарастает.
Двойные (цис-) связи в жирнокислотных ненасыщенных цепях мембран могут играть роль зародышей образования кинков в соседних насыщенных цепях. В этом случае для образования кинка в ненасыщенной цепи необходимо появление лишь одной гош-конформации при искривлении цепи на 80°. При этом устраняются стереохимические затруднения, возникающие при размещении ненасыщенной цепи в углеводородной зоне мембран из насыщенных липидов. Это хорошо согласуется с экспериментально наблюдаемым резким снижением температуры фазового перехода мембран из насыщенных липидов при добавлении к ним небольших количеств ненасыщенных жирнокислотных цепей – мембрана становится более рыхлой. Поворот вокруг С − С-связи с образованием транс-конформации практически исключен.








