 2014-02-12
2014-02-12 1395
1395Активный транспорт кальция
Са2+-зависимая АТФаза, сопряженная с кальциевым насосом, локализована в мембранах саркоплазматической сети и имеет сходные черты с К+-Na+-АТФазой. Са2+-зависимая АТФаза состоит из одной полипептидной цепи с молекулярной массой около 100 000 и относительно высоким содержанием гидрофобных аминокислот. Для работы Са2+-зависимая АТФазы необходимо также присутствие фосфолипидов.
Схема работы Са2+-зависимая АТФазы может быть представлена следующим образом. На первом этапе происходит связывание Са2+ и АТФ. На втором этапе АТФ гидролизуется с образованием фосфорилированного фермента. Образующаяся фосфорилированная форма фермента [E ~ P]* конформационно неустойчива и претерпевает изменения таким образом, что ион-связывающие участки оказываются отделенными от внешней среды.
На следующем этапе цикла происходит изменение сродства Са2+-связывающих центров к ионам кальция одновременно с изменением характера связи фосфатной группы и фермента. Энергия, ранее сосредоточенная в связи комплекса [E ~ P]*, расходуется на изменение константы связывания ионов кальция с ферментом. Вследствие изменения пространственной структуры фермента ионы кальция получают доступ во внутреннее пространство мембранных пузырьков и выбрасываются во внутренний объем. Изменение свободной энергии при этом составляет ∆G = 17,8 кДж/моль. Таким образом, суммарные затраты энергии на перенос Са2+ через мембрану примерно вдвое меньше энергии гидролиза АТФ (∆G = − 36,0 кДж/моль). Этого хватает на перенос двух ионов кальция.
Активный транспорт ионов кальция широко представлен в мышечных клетках.
В функционировании биоэнергетических систем важное место занимает транспорт протонов. Перенос электронов в мембранах митохондрий, хлоропластов и бактерий сопровождается трансмембранным переносом катионов водорода Н+ и образованием градиента электрохимического потенциала.
Перенос Н+ через мембраны может осуществляться механизмами трех типов:
- в некоторых мембранах существуют подвижные переносчики протонов (пластохинон в фотосинтетической мембране хлоропластов);
- возможен также конформационный переход мембранного белка при связывании протона на одной стороне мембраны и депротонировании белка с другой стороны мембраны, сопряженные с поворотом макромолекулы в мембране, при котором присоединенный протон пересекает мембрану;
- протоны могут транспортироваться через мембрану по специализированным Н+-каналам.
В биологических мембранах системы водородных связей, по которым транспортируются протоны, образованы полярными группами, не участвующими в формировании α-спиралей полипептидной цепи. Эстафетная передача возможна лишь при чередовании стадий дефекта (рис.5.12).
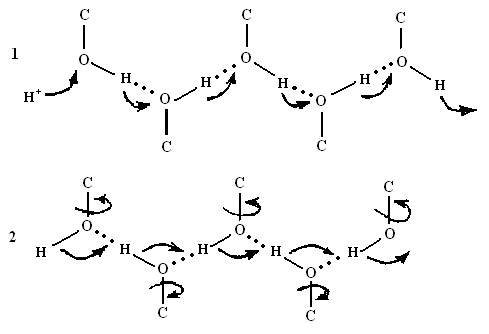
Рис.5.12.Проведение протона по цепи водородных связей (1 – стадия перескока протона; 2 – стадия поворота групп или миграция поворотного дефекта).
Она возможна лишь при чередовании стадий перескока протона и поворотов химических связей в области образовавшегося дефекта. Транслокация одного протона протекает в две стадии. Сначала происходит перескок протона от группы к группе, а затем поворот группы, в результате которого проходит дополнительное смещение заряда, и система связей возвращается в исходное состояние. В результате поворота химической группы или молекулы воды образуется конфигурация, не допускающая возврата протона в начальное положение. В жесткой структуре α-спирали повороты групп затруднены, поэтому система Н-связей α-спирали не способна проводить протоны.
В общем виде систему активного транспорта рассматривают как трансмембранный канал с большим числом участков связывания Н+, в котором транслокация протонов происходит за счет последовательных перескоков Н+ между участками связывания. Такой канал может работать как ионный насос, если структура энергетических барьеров претерпевает динамические изменения за счет энергообеспечивающей реакции. В этом случае протон будет освобождаться преимущественно с одной стороны мембраны, а поглощаться – с другой.

