 2015-02-18
2015-02-18 8835
8835Қазіргі уақытта өндірісте қолданылатын фермент препараттарының бағасы қымбат, ал өндірістік циклде бірден ерігіштігіне байланысты қолданылады. Бұл жағдай периодты процестерді үздіксіз технологиялық режимге ауыстыруға, сондай-ақ ферменттативті реакцияны басқаруға мүмкіндік бермейді. Фермент препараттарының құрылысының қазіргі заманғы білімі, оның активтілік орталығының құрамы, функциялық топтың мәні каталитикалық активтілігінің байқалуы ферменттердің пролонгирлеуші әсеріне немесе иммобилизденген фермент препараттарын құруға мүмкіндік жасады. Фермент иммобилизациясының негізі оларды ерімейтін негізге бекіту немесе жартылай өткізгіш мембрана қабықшаға енгізу.
Иммобилизденген фермент дегеніміз ерімейтін биокатализаторлар, оларда ферменттер химиялық немесе физикалық (адсорбция күшімен) түрде қандайда бір тасмалдағышпен байланысқан, матрицаға немесе микрокапсулаға бекітілген. Ферментті тасмалдағыштар немесе матрицалар мүлдем ерімейтін, химиялық және биологиялық тұрақтылығы мен ерекшеленетін, механикалық беріктігі жоғары болуы тиіс. Иммобилизделген ферменттерді алу үшін полимерлі тасымалдағыш негізіндегі табиғи (целлюлоза және оның туындылары) және жасанды (полистирол, полиакриламид, полиамид) полимерлер кеңінен қолданылады.Тасымалдағыш ретінде сонымен қатар саңылауы бар шыныны, сазды, силикагельді, мата, қағаз және т.б. материалдарды қолданады.
Ферменттер иммобилизациясы өте қиын мақсатта және әрбір фермент үшін жеке–жеке есептелінеді. Ферменттерді иммобилизациялаудың химиялық және физикалық әдістері бар. Физикалық әдістер ақуыздардың әртүрлі беттерде адсорбцияланатын қабілетіне негізделген. Адсорбция әдісі өте қарапайым, бірақ оның кемшілігі болып, иммобилизделген ферменттердің оңай десорбциясына байланысты.
Фермент тасымалдағышпен өзара әрекетке түспейтін иммобилизациялық әдісі бар. Онда фермент гель торымен ұяшықтарына енгізілген болуы мүмкін.Гель түтікшесінің үлкендігі фермент молекуласының көлемінен кіші болуы тиіс, бірақ олар субстраттың ферментке әсер етуіне кедергі жасамауы тиіс. Фермент иммобилизациясы үшін акриламид гелі кеңінен қолданылады. Бұл әдіс субстрат жоғарғы молекулалы заттар болғанда жарамсыз болады.
Ферменттерді иммобилизациялаудың химиялық әдісі матрица мен ақуыз молекуласының арасында ковалентті байланыстың түзілуіне негізделген. Химиялық әдістің артықшылығы пролонгирлеуші әрекеті биокатализаторларды алуға мүмкіндік береді: фермент ұзақ уақыт бойы қолданылса да ерітіндіге өтпейді.
Фермент иммобилизациясымен қатар микроорганизмдердің бүтін жасушасы да иммобилизациялануы мүмкін. Бұл әдіс полимер кеңістігінің торына жасушаның механикалық негізделуіне негізделген, онда олар өзінің тіршілік етуіне және ферменттік активтілігін сақтайды. Иммобилизация кезінде тәртіп бойынша фермент активтілігі төмендейді. Иммобилизденген формада фермент активтілігі бастапқы еритін ферменттің активтілігінің 10-90% құрайды. Осыған қарамастан, өндірісте иммобилизденген ферментті қолданған айтарлықтай тиімді.
Иммобилизация көп жағдайларда ферменттердің тұрақтылығына әкеп соғады. Реакторда иммобилизденген ферменттер бір апта немесе бір ай көлемінде қолданылады, демек бұл еритін формадағы ферменттерді қолданумен салыстырғанда экономика жағынан эффективтілігі жоғары болады.
Тест сұрақтар
1.Технологиялық сызба нұсқадағы беттік культивирлеуде болмайтын қондырғы:
А) биореактор;
В) өсімдік камерасы;
С) кюветалар;
Д) залалсыздардырғыш;
Е) дұрыс жауап жоқ.;
2. Фермент продуцентін беттік культивирлеуде... қолданылатын:
А) көлемді аэрация тәсілі;
В) минералды тұздарды қосмша қосу тәсілі;
С) СоС12;
Д) Н2 SO4;
Е) мырыш сульфаты;
13 – сабақ. Полисахарид, липид алу
Сабақ жүргізу формасы: лекция –конференция
Дәріс жоспары:
1.Липид туралы қысқаша мәлімет
2.Липид өндірісі
3.Полисахаридтер өндірісі
Липид туралы қысқаша мәлімет
Липидтер дегеніміз – бос май қышқылдары мен спирттердің күрделі эфирлері. Олар суда нашар ериді немесе ерімейді, ал хлороформ, гексан, метанол, бензин сияқты органикалық ерітінділерде өте жақсы ериді. Ақуыздармен, көмірсулармен қатар липидтер де жануарлар, адамдар және өсімдіктердің барлық ұлпаларының жасушаларының құрамына кіреді. Липид – гректің lipos – май деген сөзінен шыққан.
Молекуласындағы май қышқылдарының санына байланысты моно-, ди- және триацилглицеридтер түзілді. Егер күрделі эфир глицериннің бір гидрооксильдік тобынан құралса, ол – моноацилглицерид. Оның құрамына май қышқылының екі молекуласы кірсе, диацилглицерид түзіледі, сол сияқты егер глицериннің үш гидроксильдік тобының үшеуі де әсерлессе, онда триацилглицерид құралады. Табиғи майларда негізінен триацилглицеридтер ғана кездеседі.
Триацилглицеридтердің атауы олардың молекулаларының құрамына кіретін май қышқылдары атауынан шығады. Жоғарыда келтірілген триацилглицерид құрамына стеарин қышқылы (бірінші жағдайда), олеин қышқылы (екінші жағдайда) және май қышқылы (үшінші жағдайда) кірген.
Липидтердің жіктелуі: Химиялық құрамы мен құрылымы жөнінен липидтер 2 класқа жіктеледі.
А. Қарапайым липидтер: май қышқылдары мен әртүрлі спирттердің күрделі эфирі.
1. Глицеридтер (ацилглицериндер) май қышқылдары мен үш атомды спирт глицериннің күрделі эфирі.
2. Балауыз (воск) май қышқылдары мен бір немесе екі атомды спирттердің күрделі эфирлері.
Жоғары молекулалы май қышқылдарының және С16 – С30 бір атомдық жоғары спирттердің күрделі эфирлері балауыздың негізін құрайды. Балауыздың құрамында бос май қышқылдары, спирттер, көмірсулар, бояушы және иісті заттар болады. Оның құрамы күрделі.
Іс жүзінде ара балының балауызы, спермецент (кашалот китінің бас қуысынан алынады), ленолин (қойдың тері майынан алынады). Ара балы балауызы құрамының негізін күрделі эфир – пальмитомирицил құрайды.
В. Күрделі липидтер:
1. Фосфолипидтер. Құрамында май қышқылдары мен спирттерден басқа, фосфор қышқылының қалдығынан тұратын қосылыс.
2. Гликолипидтер.
3. Стероидтар.
С. Липидтердің туындылары
Қарапайым липидтер мен күрделі липидтердің ыдырауы кезінде түзілген аралық өнімдердің ішінде гидрофобтық қасиет көрсететін барлық қосылыстар, атап айтқанда қаныққан және қанықпаған май қышқылдары, жоғары дәрежелі спирттер, холестерин және оның туындылары (стероид гормондар, өт қышқылдары, витамин Д), көп қанықпаған май қышқылдарының (КҚМҚ) кейбір биологиялық активті туындылары, мысалы, простогландиндер, тромбоксандар, лейкотриендер және липоксиндер осы топқа жатады.
Сонымен қатар тағам құрамында міндетті түрде көп қанықпаған май қышқылдары болуы керек. Олар организмде синтезделмейді. Демек, бұл май қышқылдары өсімдік текті тағамдар немесе теңіз жануарларынан жасалған тағамдар арқылы түсіп отыруы қажет. Сол себепті оларды алмастырылмайтын май қышқылдары немесе эссенциале деп атайды. Витамин Ғ деп те атайды. Оларға линол С17Н31СООН 18:2, линолен С17Н29СООН 18:3, арахидон қышқылдары С19Н31СООН 20:4 жатады.
Мал организміндегі май тері асты шелінде, шарбыда, шыжырқайда, қойдың құйрығында, түйенің өркешінде орналасады. Өсімдік организміндегі май тұқымында (дәнінде), сабағында, тамырында болады. Азықтық майдың көп бөлігін мақта, күнбағыс дәнегінен, соя мен жүгері дәнінен алады. Сол сияқты сүттен және мал етінен де май алынады.
Мал мүшелері мен ұлпасындағы липид мөлшері әр түрлі. Мысалы, бұлшық етте 2% шамасындай май, 0,5 % фосфолипидтер, 0,1 % стеролдар бар. Майлы ұлпада (шарбыда, тері асты шелінде) майдың мөлшері 50 % және одан да көбірек.
Майлардың химиялық қасиеттері
Триацилглицеридтер (ТАГ) химиялық тұрғыдан бейтарап, инертті болып келеді. ТАГ-ге негізінен мынадай реакциялар тән:
Гидролиз реакциясы. Бұл реакция әсіресе майларға тән нәрсе. Тірі организмде гидролиз реакциясы катализатор (фермент) әсерімен жүреді, ал организмнен тыс жерде сілтілік катализатор қызметін атқарады және қыздыру қажет етеді. Егер майдың гидролизденуі сілтінің әсер етуімен жүретін болса, ондай реакцияны сабындану деп атайды. Бұл кезде май қышқылдарының тұзы түзіледі, ондай тұздар сабын деп аталады. Сабын дегеніміз – май қышқылдары натрий тұздарының қоспасы. Мысалы, С17Н35СООNa, C19H33COONa негізінен тұздарға жатады. Май қышқылдарының калий тұздары сұйық болады, олар медициналық сабын деп аталады. Кальций тұздарының жуып тазартатын сабындық қасиеті нашар.
1. Гидрогендену реакциясы. Әдетте мұндай реакция сұйық майдан қатты май алу мақсатымен іске асырылады. Маргарин өндірісінде саломас алу үшін қолданылады.
2. Гидроасқын тотықтардың түзілу реакциясы. Бұл реакция ферменттердің қатынасынсыз өтеді. Липидтердің қос байланысы бар май қышқылдарын күшті тотықтырғыштар тотықтырады. Мысалы, оттегінің асқын тотығы Н2О2, супертотық-анион радикал О2 немесе гидроксил-радикал ОН. Мұндай тотықтырғыштар жасушада болады.
Қанықпаған май қышқылдары тотығу процесі кезінде гидропертотыққа айналады:
 С=C + Н2О2 H - O - O - C – C
С=C + Н2О2 H - O - O - C – C
Қанықпаған тотықтырғыш гидроасқын тотық
қышқылдың
фрагменті
Мембрана құрамына кіретін липидтердің қанықпаған қышқылдары оңай асқын тотық түзеді, ондай тотықтар – улы қосылыстар. Липидтердің асқын тотығы жасушаға зиянды әсер етеді, мембраналық белоктарды тотықтырады, оның салдарынан мембрана белоктарының құрылымы мен қызметі өзгереді.
Тотығып ашуға қарсы әр түрлі заттар (токоферол (Е витамині), ретинол (А витамині) және аскорбин қышқылы (С витаминдері)), каротиноидтар, глутатион) қанықпаған қосылыстарды асқын тотығып ашудан сақтайды.
3. Майлардың асқын тотығып ашуы, бұзылуы.
Триацилглицериді ұзақ сақтау кезінде және қанықпаған қышқылдар тотығуының жағымсыз кезінде оттегінің әсерінен гидроасқын тотықтар ғана түзіліп қоймайды, сонымен қатар мынадай құрылымды асқын тотықтар да түзіледі:
 - СН=CH - + O2 - CОH - CОH –
- СН=CH - + O2 - CОH - CОH –
қанықпаған қышқыл молекуласының асқын тотық
фрагменті
Асқын тотықтар әрі қарай тотығуы мүмкін, бұл кезде -С-С- арасындағы коваленттік байланыс үзіледі де, альдегидтер, кетондар, төмен молекулалы карбон қышқылдары сияқты тотыға түскен қосылыстар түзеді. Тотығудың мұндай өнімдері азық-түлік өнімдерінің сапасын нашарлатады, майды ашытып бүлдіреді.
5. Майдың «кебуі». Өсімдік майлары, әсіресе зығыр майы қыздыру кезінде қоюланады да бара-бара қатайып, түссіз жарғақ секілденіп қалады. Бұл қанықпаған қышқылдардың полимерленуіне, сөйтіп полимер түзілуге және оттегін қосып алуға байланысты мәселе. Соның нәтижесінде О2 есебінен майдың массасы ұлғаяды. Осындай жолмен олифа майлы бояулар, лактар алынады.
Майлардың физикалық қасиеттері
Майлардың физикалық және химиялық қасиеттері құрамындағы қышқылдарына байланысты. Мал майы (сиыр майы, сүт майы және басқалар) 40-60 % қаныққан май қышқылдарынан (негізінен миристин, пальмитин және стеарин қышқылы), 30-35 % моноқанықпаған май қышқылдарынан тұрады. Мал майында полиқанықпаған май қышқылдары аз. Сондықтан мал майы үй температурасында қатып қалады. Жылқы майы мен қаз майында полиқанықпаған май қышқылдары көп. Сондықтан да олардың майы қатпайды.
Өсімдік майында әдетте қаныққан май қышқылдары аз (10-20 %), қанықпаған қышқылдары көп (80-90 %). Қанықпаған қышқылдардың құрамы өсімдік түріне байланысты болады. Зәйтүн майының 79 % олеин қышқылынан, күнбағыс майының 75% линоль қышқылынан тұрады. Өсімдік майлары сұйық күйде болады.
Майдың азық - түлік ретіндегі бағалылығы оның балқу температурасына байланысты. Майдың балқу температурасы мал денесінің температурасына жақындаған сайын, май жақсы эмульсияланады, асқазанда жақсы қорытылып, организмге жақсы сіңеді.
Май тәріздес заттар (липоидтер)
Май тәріздес заттарды кейде күрделі липидтер деп атайды. Оларды келесі топтарға бөледі:
1) фосфолипидтер (фосфотидтер),
2) гликолипидтер,
3) стеролдар (стериндер) мен стеридтер.
Липидтердің алмасуы. Липидтер организмге түскеннен кейін ас қорыту жолдарында сәйкес ферменттер эстеразалардың әсер етуімен құрамдас бөліктерге ыдырайды. Ас түйірі ауыз қуысына қарапайым механикалық өңдеуден өтіп асқазанға түседі, себебі сілекейде майларды ыдырататын ферменттер жоқ.
Майлардың асқазанда қорытылуы. Майдың қорытылуы асқазанда басталады. Асқазанның қозғала алатын ерекшелігіне байланысты, майдан қою эмульсия түзіледі. Бұл құбылыс ферменттердің әсерін жеңілдетеді. Қарындағы қоректердің ішінде липаза бар. Бірақ оның қасиеттері ұйқы безі липазасының ең қолайлы рН деңгейі 6-7 болса, қарын липазасы рН көрсеткіші 3,5 болғанда активті. Бұл липаза көбінесе көміртегі атомдарының тізбегі ұзын емес, орташа болатын май қышқылдары эфирлерін гидролиздейді.
Бұл кезде босап шыққан май қышқылдары тікелей қарын арқылы сіңеді. Жаңа туған сүтқоректілерде липаза ерекше қызмет атқарады. Өйткені сүтте төменгі молекулалық май қышқылдарының триацилглициридтері болады.
Майларды алу. Майларды тек табғи көздерден ғана алуға болады. Жануарлардың ұлпасынан майды қайырып бөліп алады. Ал өсімдіктер майын сығымдап немесе органикалық еріткіштермен (хлороформмен, бензолмен немесе эфирмен) экстракция жасау жолымен алынады.
Май адамға және малға қоректік өнім ретінде қажет. Олардың құрамында алмастыруға келмейтін май қышқылдары бар. Биологиялық маңызы жөнінен май ақуыздан кем емес. Май азықтық мақсаттан басқа техникада майлау ретінде қажет, өнеркәсіпте майлардан әр түрлі сабын, бояу, олиф сияқты заттар дайындалады.
Липид өндірісі
Қазіргі кезде азықтық мақсатында өсімдік және жануарлар организмдерінен алынатын липидтердің және майлардың санаулы көлемі техникалық қажеттілікке, тамаққа жұмсалады. Липидтерді алудың жаңа түрі микроорганизмдердің кейбір түрлері азықтық емес шикізатта липидтің белгілі мөлшерін жинап алады. Мұндағы липидтер деп отырғанымыз полярлы емес еріткіштерде еритін микоорганизмдердің жасушасының компоненті. Микроорганизмдердің липидтері (биожир немесе микробтық май) әр түрлі халық шаруашылығында (медицинада, химия-фармацевтикада, металлургияда, лак бояуларда және тағы басқа жерлерде) қолданылуы мүмкін. Оның осы қасиеттері азықтық мақсаттағы жануар мен өсімдік майының белгілі мөлшерін үнемдеп алмастыра алады.
Микроорганизмдерді әр түрлі субстратта культивирлеу процесінде қарапайым липидтерді (бейтарап майлар), күрделі липидтерді (фосфолипидтер және гликолипидтерді) және липидтің туындылырын (май қышқылы, спирттер, көмірсулар және витаминдер) алуға болады.Жалпы липидтер микроорганизмнен басқа барлық жерде қатысады, бірақ та кейбіреулері липидтің белгілі мөлшерде жиналу қасиетіне ие болады.

Липидтердің микроорганизм - продуценті болып ашытқылар Candida Lipomyces, микроскоптық саңырауқұлақтар Rhizopus, Fusariun туыстары және бактериялар Mycobacterium Lactobacillus саналады.
Жеке микроорганизм - продуценті 60-75 % липид түзе алады(есеп бойынша құрғақ биомасса).Микроорганизмдердің әр түрлі систематикалық топтарын культивирлеу барысында алынған липидтердің құрамы өзгеше болатындығы байқалады, оны липид түзуші микроорганизмдерді таңдауда ескереді.Мицелиалы саңырауқұлақтардан Penicillum, Rhizopus, Fusariun 40 -70 %- ке дейін липид жинайды.Бактериялардан микробактерии 40 %- ке дейін липид жинайды.Бірқатар бактериялар полигидроксибутират саны 60 %- ке дейін жетеді,мысалы водород тотықтырғыш Alcaligenes eutrophus түрі. Белгілі культивирлеу жағдайда 60 %- ке дейін және кейбір балдырлардың микроформаларында липид көбірек жинайды
Қоректік орта құрамы. Липидтің микроорганизм- продуценті үшін көміртегі көзі және энергия көзі болып көмірсулар, органикалық қышқылдар, көмірсутектер, спирттер болып табылады.Сондықтан микробтық липид алуға арналған шикізат болып, азықтық ақуыз алу өндірісінде қолданылатындар саналады: ағаш қалдығының гидролизаты, азық – түлік өндірісінің қалдығы, көмірсутегі қолданады.
Көміртегі көзі липид түзу процесіне ғана емес, сондай-ақ оның құрамына да әсер етеді. Липид синтезіне қоректік ортадағы көміртегі мен азот қосылысы да айтарлықтай әсер етеді. Азот концентрациясының бірден жоғарылап кетуі микроорганизмдердің липид түзу қасиетін төмендетеді,
ал азот көзінің жетіспеуі клеткада липидтердің құрамының көбейіп кетуіне әкеліп соғады. Микроорганизмнің өсуі және липид түзуші процестің жылдамдығы, сондай-ақ минералды азық көзіне де байланысты. Микроорганизмдердің өсуі және липидтердің жиналуына минералды тұздардың ішіндегі ең көп әсер ететіні фосфат болып табылады, фосфордың артық болуы ақуыз синтезінің жоғарылауына әкеліп соғады,ал фосфордың жетіспеушілігі ортада липидтің түзілуін арттырады.
Липид түзуші микроорганизмдердің көпшілігі прототрофты болып табылады, демек әрбір витаминнің продуценті, сондықтан олар құрамында осы қосылыстар жоқ ортада өсе алады. Бірақ қоректік ортаға кейбір витаминдерді қосқанда липидтің түзілуіне және оның құрамына қолайлы әсер етеді.
Культивирлеу әдісі. Липидті продуцирлеуші – аэробты микроорганизмдерді өсіру процесіне ортаның рН және температурасы сияқты факторлар әсер етеді. Липид түзілу процесіне ерітілген оттегі концентрациясының әсері туралы әзірше нақтылы пікір жоқ, бірақ та оттегі липид биосинтезіне қолайлы әсер етеді деген болжам бар. Ферментация процесі кезіндегі аэрациялаудың қарқындылығының өзгеруі липид құрамына әсерін тигізеді.
Микроорганизмдердің өсу жылдамдығы мен липидтің пайда болуы рН ортаның әсеріне байланысты, әр продуценттің өзінің рН ортасы қолайлы болады. рН ортаның қолайлы жағдайынан ауытқуы, липид өндірілген өнімнің көлемінен гөрі оның құрамына қатты әсер етеді. Көптеген микроорганизмдердің өсуі үшін температураның қолайлылығы және липидтің пайда болуы сәйкес болы керек. Микроорганизмнің жасушасындағы жалпы липидттің мөлшері культивирлеу температурасына онша байланысты болмайды. Температураның өзгеруінің әсері продуценттің өсуінің жылдамдығына және биомассаның жиналуына әсер етеді, ал липидтің құрамына мүлдем әсер етпейді. Липидтің құрамы культивирлеу температурасына байланысты шамалы өзгеріске ұшырайды. Осыған байланысты қандай да бір дәрежеде температураның өзгергенінен липид фракциясының құрамын реттеп отырады.
Липид өндірісінің технологиясы. Қазіргі кезде микробтық липид алуда ең көп өңделген технологиялық режим гидролизат трофында және көмірсутекте ашытқыларды өсіру болып табылады. Бірақ өндірістік масштабта арнайы микроорганизмдердің штамын культивирлеу арқылы үлкен көлемде липидті синтездеу әдісі әлі құрылған жоқ.
|
| ||
| ||
| ||
|
Гексан
 | |||
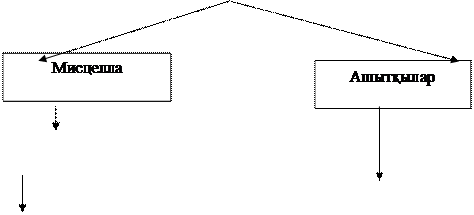 |
Гексан
 | |||
| |||
|
Бу Гексан
 |  | ||||||
|  | ||||||
|
61 сурет - Липид өндірісінің технологиялық сызба нұсқасы
Мұнай парафинінен Candida туысындағы азықтық ашытқыдан алынатын липид маңызды. Ашытқы жасушасында липидтердің құрамы 16-20% жоғары, басқа липид түзушіге микроорганизмге қарағанда (құрғақ биомасса есебі бойынша) жоғары болады.
Демек, липидтерді ашытқылардан бөліп алу экономика жағынан тиімді болады, өйткені үлкен көлемде азықтық ашытқы өндіріледі, ал липидтер, бұл азықтық қоспаға қажетсіз компонент болып табылады.
Липид өндірісі кезінде азықтық ашытқыны алудың технологиялық сызба нұсқасында соңғы өнімді бөліп алу сатысына тағы бір – құрғақ ашытқыдан органикалық ерітінді арқылы липидті экстракциялау операциясы қосылды. Осы технологияны орындағаннан соң екі өнім – «майсыздалған» ақуыз концентраты және микробтық май алынады.
Азықтық ашытқыны алу технологиясы ертеректе қарастырылған, сондықтан липидті азықтық ашытқыдан бөліп алу сызба нұсқасын қарастырамыз.
Шашыратқыш кептіргіштен алынған ашытқыны арнайы ыдысқа салып ылғалдайды, содан соң қыздырып майдалап түйіршіктеуге жібереді. Ылғал биомассаны майдалау фильтер арқылы қысып жаншып үгітетін машинаға (бумен жұмыс жасайды) түсіреді.
Жаншып үгітетін машинада түйіршіктелгендер ісініп нәтижесінде жасуша бұзылады; алынған өнім пневматикалық кептрігішке тасымалданады. Кептіргіш тігінен орнатылған құбыр болып келеді, оның бойымен ісінген, майдаланған түйіршіктер және қыздырылған кептірілген агент (ауа) жоғары тура ағыммен жылжиды.
Кептірілген биомасса түйіршіктері майдалануға түсіп, онда жұқа фракцияға бөлінеді (түйіршік бөлшектің размері 2 мм кем емес), ал негізгі масса экстракцияға жіберіледі.
Экстракция құрылысы әр түрлі экстракторларда қатты заттарды экстракциялау принципі бойынша жүргізіледі. Экстрагент ретінде органикалық еріткіштерді қолдануға болады. Экстракциялау нәтижесінде алынған биомай ерітінідісін еріткіштегі мисцелла деп аталады. Биомай мисцелланы бөліп алу үшін дистиляцияға еріткішті айдау үшін жібереді.
Дистилляция жүйесі әдетте екі сатыдан тұрады. Дистилляцияның бірінші сатысында еріткішті айдау процесі биомай концентрациясын 90%-ке жеткенше қабықшалы - буландырғышта жүргізеді. Еріткішті айдаудың соңғы сатысы вакууммен дистилляциялы бағанада жүргізеді.
Экстракциядан кейін құрамында аздап еріткіші бар майдан тазартылған биомасса арнайы десольвентор деп аталатын өткір бу берілетін аппаратқа келіп түсіреді. Бу мен жоғарғы температураның әсерінен ашытқы еріткіштен ажыратылады және кептіріледі. Дайын биомасса десольветордан түйіршіктеуге келіп түседі және кептіруге жібереді.
Микробтық синтез липидті қажет ететін халық шаруашылығының әртүрлі салаларын қанағаттандыра алады.
Полисахарид өндірісі
Полисахаридтер немесе гликандар –11 моносахаридтер бірлігінен кем емес полимерлерден құрастырылған.Олар бір немесе бірнеше моносахарид түрлерінен тұрады. Гомополисахаридтер және гетерополисахаридтер өзінің құрылысына қарай ажыратылады. Полисахаридтер– барлық организмдегі міндетті түрде болатын қажет компоненттердің табиғатта кездесетін көмірсулардың үлкен бөлігін құрайды демек, жер шарындағы органикалық заттардың негізгі массасы өсімдік биомассасында басым болады.
Полисахаридтер полимерлер түрінде кездеседі, сондай-ақ нуклеин қышқылымен, ақуызбен, липидпен, фосфатпен комплекс түрінде кездеседі. Мономер құрамы мен құрылысы әртүрлі.Полисахаридтің микроорганизмдері әртүрлі болып ажыратылады. Оның кейбіреулері өсімдік пен жануарлар полисахаридіне жақын немесе ұқсас бірдей болып келеді. Микробтық полисахаридтердің басым көпшілігі түріне арналған немесе серологиялық тобының түрі үшін арнайы құрылысқа ие. Микробтық гликандар ертеректе белгісіз болған моносахаридтерде жиі кездеседі, олар жануарларда да, өсімдіктерде де кездеспейді.
Соның ішінде көптеген микроорганизммен пайда болған слизь Пастердің кезінде білгілі болған көмірсу табиғатына ие болады. Бірақ микробтық полисахаридтерге өткен 100 жылдықтың 20 жылынан бастап көңіл бөліне бастады, пневококканың серологиялық ерекшелігін анықтаушы деп полисахарид саналды. Қазіргі кезде халық шаруашылығында және медицинада кеңінен қолданылатын мүмкіншілігіне қарай, микробтық полисахаридті зерттеуде ерекше мәнге ие болып отыр.
Микроорганизмдердің полисахаридтері клетканың ішіндегі және клетканың сыртындағы болып локализацияға сәйкес бөлінеді. Клетка ішіндегі полисахаридтер цитоплазмалар, мембрана және клетканың қабырғасы жатады. К летка сыртындағы – полисахаридтерге капсула, чехли ( қабырға құрылысы) және бос слиз клетка қабырғасына жатпайтындар саналады. Кейде клетка сыртындағы полисахаридтерге, цитоплазмалық мембрана сыртынан локализделгендер жатады. Мұндай жағдайда клетка сыртындағы және клетка қабырғасының полисахаридтерінің тобына жатады. Бірқатар микроорганизмдерде капсуламен клетка қабырғасының бір-бірімен шекарасын ажырату қиын.
Локализациялауда полисахаридтер үш топқа бөлінеді: клетка ішіндегі (цитоплазмалар, мембрана, периплазмалар), клетка қабырғасындағы полисахаридтер және клетка сыртындағы (капсула, чехли және бос слиздер) болып бөлінеді.
«Экзогликандар» термині негізінде бос слизді полисахаридте қолданылады. Кейде капсулалы полисахаридтерді экзогликан деп атайды.
Микробтық полисахаридтер топтарға біріктіріледі және функциясы бойынша: активті тасымалдағышқа қатысушы қорда тұрған, қолдаушы, клетканың бір-бірімен әсерлесуіне қатысушы, қорғаушы және т.б.
Кейбір зерттеушілер оның топологиясына және функциясына қарай жіктейді. Клеткалық полисахаридтер осыған сәйкес екі топқа бөлінеді. Біреуі резервті-энергетикалық және модификаторлар қосады (клетка ішіндегі), екіншісі – құрылыстық және құрылысты – метаболиттік (клетка қабырғасында) болып бөлінеді. Клетка сыртындағыға құрылысты – метаболиттік гликандар гиперөнімді және экзогликандарға қабілетті болып саналады.
Цитоплазмалардың және мембраналық құрылысының полисахаридтері
Цитоплазма полисахаридтері екі формада пайда болады: олар оған диспергирленген болуы мүмкін немесе түйіршіктері бір-бірімен біріктірілген болуы мүмкін. Бактерия цитоплазмасы 20-30 % полисахаридтен тұрады, клетканың құрғақ массасынан 50-60 % дейін жиналу қабілеті бар. Көбінесе микроорганизмдердің цитоплазмасында гомогликандар болады, одан әсіресе гликоген типті глюкандар таралады. Оларды прокариотты және эукариотты микроорганизмнің цитоплазмасынан бөліп алады: әтүрлі бактерияның өкілдерінен Agrobacterium, Arthrobacter, Bacillus, Clostridium, Esherichia, Mycobacterium, Nostoc, Rhodoрseudomonas, Rhodospirillum, Streрtococcus, сонымен қатар, ашытқыдан, мицелиалы саңырауқұлақтан, қарапайымнан, балдырлардан бөліп алды.
Гликоген сияқты полисахаридтерден басқа бірқатар микроорганизмдердің цитоплазмасында крахмал, маннан, леван, арабан және ксиландар табылған. Гликоген және тағы басқа да ДНК, РНК, ақуызбен, липидпен, фосфатпен комплекс пайда болатын цитоплазмада гомогликандар табылған. Гетерополисахаридтер цитоплазмада сирек кездеседі. Бірақта Streptomyces және Mycobacterium өкілдерінде олар басым болып асып түседі екен.
Цитоплазманың полисахаридтерінің функциясы соңына дейін анықталмаған. Қазіргі кезге дейін көміртегі көзінің қорындағы тұрған зат және клеткаға арналған энергия қызметін атқаратын зат деп санаған. Олар мысалы, Bacillus, Clostridium бактерияларының туысындағы эндоспердің жетілуінде жұмсалады. Демек, түсінікті цитоплазманың полисахаридтері бірқатар маңызды функцияны атқарады. Клетканы реттеу механизмінде, әртүрлі заттардың синтезін бақылауда және клетканы бөлуде комплекс гомогликандар басқа цитоплазманың компонентімен бірге қатысумен жүреді.Сондай-ақ гликогенрибосомалы комплекстер ақуыз синтезін бақылауға мүмкіншілігі бар.Гликоген нуклеин қышқылының молекуласымен байланысты радиоқорғағыш әсер етеді.
Микроорганизм мембранасында орташа 2-5% дейін, кейде мембрана массасынан (Micrococcus luteus- да) 15-20% дейін көмірсулар болып табылады. Кейде клетка қабырғасында немесе цитоплазмада қалдық материал болып осы көмірсу толығымен немесе бір бөлігі болады екен. Грам оң, кейде грам теріс бактериясының мембранасында гликолипидтер және гликопротеиндер болады екен.
Микрококка мен кейбір стрептококканы санамағанда, барлық грам оң эубактерия сондай – ақ ашытқы және мицелиялы саңырауқұлақ цитоплазмалы мембрана аумағында болады, тейхой қышқылы (құрғақ массадан 1-2%) болады. Тейхой қышқылы құрылысы басқаша қышқыл полисахаридке жатады. Оның гидролизі кезінде моносахаридтпен бірге басқа класс қосылысына жататын зат пайда болады. Әр түрлі тейхой қышқылы негізінде қалдық қанттың қатысқаны және әр түрлі типтік байланысының бар екені анықталған. Кейбір микроорганизмде тек липотейхой қышқылы бар екені анықталған, ал бос тейхой қышқылы болған жоқ. Грам теріс бактерияның мембранасында тейхой қышқылы болмаған.
Полисахарид биосинтезде және қант тасмалдауда мембраналы гликолипид қатысады. Тейхой қышқылы ион алмасуды реттейді, амин қышқылы тРНК байланысында әсер етеді, клетка қабырғасының мембранамен байланысымен іске асырылады, антигенді активтілігін байқалтады.
Клетка қабырғасындағы полисахаридтер
Полисахаридтердің грам оң эубактериясында жасуша қабырғасының құрғақ массасынан 30-60% дейін құрайды. Оның біраз бөлігі муреинді комплекс құрамына кіреді, клетка қабырғасының заттары грам оң эубактериядағы оның мөлшері 50-90% жетеді. Түзу сызықты полисахарид муреин тізбегі N – ацетилглюкозамин және N – ацетилмурам қышқылы β – 1,4 байланысымен тұрғызылған. D – сүт қышқылынан құралатын глюкозамин туындысы - мурам қышқылы болады.
Кейбір архебактерияның клетка қабырғасы, грам оңмен бояуымен боялатын псевдомуреинді гликан бөлігін құрайды, ал N – ацетилглюкозаминнен, N – ацетилгалактозаминнен және N – ацетилталозаминурон қышқылынан тұрады.Мурамо қышқылы псевдомуреинде табылған жоқ. Бірқатар клетка қабырғасының грам оң архебактериясы, тек қышқыл гетерополисахаридтерден тұрғызылған, оның құрамына галактозамин бейтарап қант және урон қышқылы құрамына кіреді. Клетка қабырғасында көбінесе бәсеңдететін грам оң эубактериясында микобактерия мен коринебактериядан басқа тейхои қышқылы болады. Оның мөлшері клетка қабырғасының массасынан 50-90 %-ке дейін жетеді, стрептомицетте ол 4-50% дейін ауытқып отырады. Микроорганизмде глицеринтейхои әлде рибиттейхой қышқылы болады. Бірақта Streptococcus fаecalis және бір ғана Streptomyces sp. штамда тейхои қышқылы екі түрінде де табылған.
Басқа полисахаридтер грам оң бактериясының клетка қабырғасын құрайтын үлкен әр түрлілігімен ажыратылады: көбінесе бұл гетерогликандар. Оның құрамында бейтарап моносахар аминосахар, урон қышқылы, ацетил тобы, фосфор қышқылының қалдығы табылады.
Грам оң бактериясының клеткасының қабырғасында 1-50 % дейінгі полисахаридтер құралады. Олардың ішінде мурейн комплексті полисахаридтер доминирлеуге жағдай жасамайды, демек оның мөлшері шамамен клетка қабырғасында орташа 5% болады екен. Тейхои қышқылы грам оң бактериясының жеке өкілдерінде табылады.
Липополисахаридтердің грам оң бактериясының клетка қабырғасына арналған – биологиялық активтік заттар ерекше сипаттайды, олар мембрананың сыртын формалауда қатысады. Әр түрлі липополисахаридтер саны көп, шапшаң жоғарғы дәрежеде оқып ізденудің өзінде, мұның көбісінің құрылысы және құрамы соңына дейін зерттелген. Комплекстің полисахарид бөлігі, базисті құрылысы және О – арнайы бүйірлі тізбегі ажыратылады. Липополисахаридтің көмірсу комплексі – бұл гетерополисахарид. Әр түрлі бактерияда оның құрамында бейтарап қант, аминосахар, урон қышқылы, метиль және ацетиль топтары болады және әсіресе 3,6-дидезокситуынды қантты, табиғатта сирек кездесетін құрамында табылады. Оның ішінде кең таралғаны абеквоза (3,6-дидезокси-D-галактоза), колитоза (3,6-дидезокси-L-галактоза), тивелоза (3,6-дидезокси-D-манноза), аскарилоза (3,6-дидезокси-L-манноза) және паратоза (3,6-дидезокси-D-глюкоза). Бұл қанттар көбінесе серологиялық арнайы бактерияны анықтайды. Моносахарид ерекше тип – гликолактилді қышқылдар О – антигенді полисахаридтер Shigella туысы өкілінің құрамында табылған. Липополисахарид (ЛПС) полисахарид комплексі бірқатар бактерия күрделілігімен ажыратылады. Олар алты және одан да көп араласқан және араласпаған моносахаридті құрайды.
Полисахаридтер – ашытқының және мицелиялы саңырауқұлақтың клетка қабырғасының басты компоненттері. Олар (Saccharomyces cerevisiae) клетка қабырғасының массасының 90% құрайды. Ашытқыда көбінесе гомогликандар табылады: гликандар, маннандар, хитин (полимер N-ацетилглюкозамин) болады. Ашытқы маннаны клетканың фосфор қалдығын құрайды және (немесе) метилді топтар, ал глюканда – ацетилді және аминді топтар болады. Көптеген ашытқының клетка қабырғасында гетерополисахаридтер – галактоманнандар және глюкоманандар, сондай-ақ 3-4 мономерден тұратын қышқыл гликандар табылады. Ашытқының клетка қабырғасында бірнеше полисахаридтер қатысады. Ашытқы клеткасының қабырғасында пептидпен байланысқан гликандар болады.
Клетка қабырғасындағы негізгі полисахарид компоненті болып мицелиялы саңырауқұлақта зерттелген хитин саналады. Сондай-ақ ацетилирленген полимер ацетилирленбеген полимер глюкозамин – хитозан, глюкандар (кейде целлюлоза), галактандар және әр түрлі гетерополисахаридтер араласқан және араласпаған қант, уран қышқылы болады.
Клетка қабырғасының микрофибрилл құрылысы целлюлозадан көбінесе микроформ балдырлардан тұрады, ал басқа гомополисахаридтің жеке өкілдері көбінесе ксилан және мананнан тұрады. Олардың саны құрғақ клетка қабырғасында 50-80% жетеді. Полисахарид матриксасы негізінде гетерогликан болып келеді.
Клетка қабырғасындағы полисахаридтер әр түрлі функцияны атқарады. Бұлардың көбісі клетка қабырғасында механикалық беріктікті көрсетеді. Сондықтан оны көбінесе «скелет» (қаңқа) деп атайды. Липополисахаридтер, тейхои қышқылы, ал сондай-ақ гетерополисахаридтер грам оң бактерия қатары клетканың антигендік активтілігіне жауапты. ЛПС біраз мөлшері грам оң бактериясы – токсинді. ЛПС энтеробактерия клетканы ингибирлеуші ұзын тізбекті май қышқылының әсер етуінен қорғайды, осы бактериялар жануарлар ішегінде өседі. Көптеген полисахаридтер литикалық ферментке және фагқа бактерияның беріктігін құрайды. Полианионді полисахаридтер клеткадан және зарядталған метаболиттерді және қоршаған ортадан затты тасымалдауға қабілетті. Одан басқада, мұндай полисахаридтер теріс зарядты клеткаға хабарлайды, нәтижесінде ортадағы клетканы бір-бірінен алшақтатады, ортада оларды диспергирлейді. Микроорганизмнің клетка қабырғасындағы полисахаридтер Н-алканды ортада өседі, жақсы эмульгатор болып саналады, клеткаға көмірсудың енуіне мүмкіндік жасайды.
Клетка сыртындағы полисахаридтер
Клетка сыртындағы полисахаридтер капсула түрінде және қаптап тұратын қап, сондай-ақ бос слиз болып келеді. Капсуланың 0,2мкм қалыңдығы жарықты микроскоппен көріп ажырату қиын, ал электронды микроскоппен көруге болады, ол микрокапсула деп аталады. Микрокапсула клетка қабырғасымен байланысты ол капсулаға қарағанда берік болып келеді. Көптеген микроорганизмдердің капсуласы белгілі құрылысы болады және слизден анық бөлінген. Кейбір бактерияда капсулалы материал бос ылжыраған болып келеді, құрылысы жоқ, клеткадан оңай ажыратылады, сондықтан капсула мен бос слизді ажырату мұндай жағдайда қиынға соғады. Мұндай аморфты капсуланы слизисті қабат деп аталады. Қабығының (чехол) капсуладан құрылысы күрделі. Мұнда әртүрлі құрылыстағы бірнеше қабатты ажырату қиын. Клетка сыртындағы полисахаридтің мөлшері клетканың биомассасын бір неше есе жоғарылатады.
Капсула, чехол және слиз барлық уақытта тек полисахаридтерден тұрмайды. Олар гликаннан басқа ақуыздарды, полипептидтерді, нуклеин қышқылын, липидтерді, полисахаридпен комплекс түзетін немесе түзбейтінді қосады. Қабығы күрделі химиялық құрамды құрайды. Демек Sphaerotilus natans глюкозаны, глюкозаминді, ақуызды, липидті және фосфатты құрайды. Бактерияның қабығы, металл қосылысты тотықтырады және тотықсыздандырады, көбінесе олардың оксидін құрайды. Кейде көмірсусыз полимерлер – капсула мен слиздің бір ғана компоненті. Капсула кейбір Bacillus туысты түрінің полипептидінен құралады, кейбір штамм Pseudomonas aeruginos слизі ДНК-дан тұрады.
Клетка сыртындағы полисахаридтер капсулалы немесе бос және басқада көптеген микроорганизмдермен құралады. Бірақ клетка сыртындағы полисахаридтер клеткада міндеті түрле атқаратын функция емес және ол белгілі бір жағдайда ғана пайда болады.
Микроорганизмнің клетка сыртындағы полисахаридтері құрылысына және құрамына қарай әртүрлі. Қазіргі кезде 200-ге жуық экзогликандар зерттелген, біріншілік құрылысы бекітілген және осының көбісінің жеке дара құрылысы бекітілген. Әртүрлі микроорганизмдердің клетка сыртындағы полисахаридінің құрамында 20-дан астам моносахаридтер және оның туындысы табылды. Көбінесе гексоза кездеседі: глюкоза, галактоза, манноза және 6-дезоксигексоза: фрукоза және рамноза. Пентоза сирек байқалады: арабиноза, ксилоза, рибоза. Урон қышқылы кең таралған: галактуронды, маннуранды және әсіресе глюкоуронды.
Көбісі аминқантын құрайды: глюкозамин, галактозамин және маннозамин. Көбінесе экзогликанда көмірсу еместер көмекшілері – пируват және ацетат, сонымен қатар сукцинат және глицерин кездеседі. Клетка сыртындағы гликан қатары үшін сирек кездесетін мономерлер бар: 2,6 немесе 3,6-дидезоксисахар болады, кейбіреулерінде бұрын анықталмаған моносахаридтер табылған, мысалы глюколактил қышқылдары (сапротрофты микробактерия). Кейде тейхои қышқылы, фосфатты және сульфатты иондар кездеседі.
Бактерияның көптеген түрінде клетка сыртындағы полисахаридтер – қышқыл гетерогликандар әртүрлі құрамда 2-5-тен, кейде 6-7 мономерден тұрғызылған. Мысалы: Xanthomonas campestric полианионит ксантан, глюкозаны, маннозаны және О-ацетил тобын және пируватты қосқатын синтездейді. Кейбір бактериялар бейтарап гетерополисахаридтерді түзеді.
Микроорганизмде әртүрлі гомополисахаридтер, әсіресе глюкандар, соның ішінде белгілісі декстриндер(бейтарап гликанға құрылысы бойынша жақын және азырақ жақын глюкон тобы) кең таралған.Олар 200000 дейін глюкозаның қалдығын құрайды, олар сызықша және тарам-тарам тәрізді болып келеді. Сызықша (негізгі) α-1,6 байланыстың қатысуымен тізбегі тұрғызылған, тарамақтысы α 1,2-, α-1,3- және α-1,4 байланыста тұрғызылған. Молекулярлық массасы декстриннің 12-600 млн аралығында ауытқуда болады. Декстриннің активті продуценті – сүт қышқылының өкілі Leuconostos mesenteroides және L.dextranicum. Декстрандар сондай-ақ кейбір Streptococcus(Str.sаnguis, Str.mutаns), Brevibacterium, Lactobacillus түрлерін синтездейді. Іс жүзінде әрбір продуцент өзінің өте жақсы декстраннен басқа түрін синтездейді. Кейбір штамдар бір мезгілде декстраннің құрылысы бойынша екі түрін түзеді.

 Клетка сыртындағы целлюлоза – полисахарид өсімдікте таралған, бактериядан кейбір өкілдері Pseudomonas, Zooglea, Azotodacter синтездейді. β- (1 3) -Глюканды курдлан деп аталатын Alcaligenes faecalis var.myxogenes және Rhizobium пайда болады.Күшті тарамдалғандар α-(1 4) -глюкан бүйіріндегі тізбегімен полимер түріндегі гликоген α-(1 6)-байланысының тізбегімен қосылған, жануарлардың полисахарид қоры, көптеген бактерияны, ашытқыны Neisseria perfiava культуральды сұйықтықтан табуға болады,Нигеран α-глюкан кезектескен (1 4) және (1 3) байланысымен Aspergillus niger саңырауқұлағы пайда болады.
Клетка сыртындағы целлюлоза – полисахарид өсімдікте таралған, бактериядан кейбір өкілдері Pseudomonas, Zooglea, Azotodacter синтездейді. β- (1 3) -Глюканды курдлан деп аталатын Alcaligenes faecalis var.myxogenes және Rhizobium пайда болады.Күшті тарамдалғандар α-(1 4) -глюкан бүйіріндегі тізбегімен полимер түріндегі гликоген α-(1 6)-байланысының тізбегімен қосылған, жануарлардың полисахарид қоры, көптеген бактерияны, ашытқыны Neisseria perfiava культуральды сұйықтықтан табуға болады,Нигеран α-глюкан кезектескен (1 4) және (1 3) байланысымен Aspergillus niger саңырауқұлағы пайда болады.
 Тарамдалған полимерлер леван –фруктозасы (2 6) байланысы сірке қышқылды бактерияны Gluconobacter oxydans, Acetobacter aceti, Pseudomonas, Erwinia тобындағы Aeromonas, Bacillus кейбір түрі синтездейді.
Тарамдалған полимерлер леван –фруктозасы (2 6) байланысы сірке қышқылды бактерияны Gluconobacter oxydans, Acetobacter aceti, Pseudomonas, Erwinia тобындағы Aeromonas, Bacillus кейбір түрі синтездейді.
 Фруктандар инсулин түріндегі (тармақталған күрделі гүлдейтін өсімдік өкілді полисахаридтер) (2 1)-байланысы негізгі тізбегімен және тарамдалған С6 Str.mutans штамы пайда болады.
Фруктандар инсулин түріндегі (тармақталған күрделі гүлдейтін өсімдік өкілді полисахаридтер) (2 1)-байланысы негізгі тізбегімен және тарамдалған С6 Str.mutans штамы пайда болады.
Маннандар Bacillus және Corynebacterium түрінің культурасында сондай-ақ көптеген ашытқыда пайда болады. Көбінесе барлық микроорганизмдер, клетка сыртындағы полисахаридтерде пайда болуына қабілетті, бос слизді және капсуланы синтездейді. Слиздің мономерлі құрамы және капсуласы көп жағдайда бірдей.
Қандай микроорганизмдер тобындағы полисахаридтер екенін анықтау барлық уақытта мүмкін емес. Бірқатар жағдайда филогенетикалық жағынан жақын бактериялар құрылысы мен құрамы жағынан бастапқы клетка сыртындағы гликандар синтездейді. Оған мысалыға бактериялы альгинат Pseudomonas aeruginosа және Azotobacter vinelаndii, курдлан Rhizobium және A.faеcalis var. myxogenes, қышқыл гетерогилкандар Corynebacterium және Arthrobacter, декстрандер Streptococcus және Lactobacillus және т.б. жатады. Бірақ та таксономикалық қатынасқа алыс тұратын микроорганизмдер гликандар пайда болады. Мұндайда полимерлер пайда болады және функциональды сәйкестікте болады. Бірақ, экзогликандар әртүрлі фитопатогенді бактериялар және минингит қоздырғыштар құрамы бойынша жақын. Бірқатар бактериялар клеткалы агрегат пайда болғанда цементтеуші материал клетка сыртындағы целлюлоза атқарады.
Белгілі бір тұқымдағы әртүрлі түрдегі микроорганизмдер көбіне және тұқымы бірдей әртүрлі штамдар бірдей емес экзополисахаридтерді синтездейді. Демек, E.coli 70 серотипке ие болады, құрылысы мен құрамы бойынша әртүрлі капсуланы полисахаридтердің және иммунохимиялық қасиеті бойынша сәйкес. Экзогликандар тиімді және түйнек бактериясының тиімді емес штамы, мономерлер құрамы бойынша, моносахарид қатынасы бойынша әртүрлі Lipomyces тұқымдас ашытқы штамы ажыратылады. Моносахарид құрамы бойынша бірдей емес клетка сыртындағы гликандар М-, S-, және R- бактерия формасы болады.
Культуральды сұйықтықта бір микроорганизм бірнеше әртүрлі гликандарды жинайды. Мысалы, L.mesenteroides декстринді және леванды түзеді, Serratia marcescens үш экзополисахаридті түзеді: қышқыл глюкорамнан, глюкурон қышқылын, рамноглюкан және гептоглюкан құрайтын Aureobаsidium (Рullularia) pullulans саңырауқұлағы екі гомогликанды түзеді: аубазидан – тармақталған полимер α-(1-4), β-(1-3) және β-(1-6) байланысы болады.
Клетка сыртындағы полисахаридтер, микроорганизмдер үшін тіршілік етуге қажет болып есептелмейді. Кез келген уақытта пайда болмайтын олардың түрлері табиғатта бар. Тәжірибе жүзінде көрсетілген клетка капсуладан (бөлінген) арылған өмір сүруге қабілетті және капсуланы жасай алады. Содан басқа клетка сыртындағы полисахаридтер белгілі функцияны орындайды, өмір сүруге қабілетті продуцент үшін қолайлы жағдай жасайды. Олардың ішіндегі біреуі барлық полисахарид үшін универсальды.
Клетка сыртындағы полисахаридтер ультракүлгін сәуледен және әртүрлі химиялық агенттен, ауыр металдан дәрілік препараттан клетканың өліп қалуынан, клетканың кеуіп кетуінен сақтайды. Капсулаланған бактерия химия терапиялық заттарға беріктеу, капсулаланбағанға қарағанда беті капсуламен қапталғаны клетканы бактерияфагтан сақтайды.
Көптеген клетка сыртындағы гликандар биологиялық жағынан активті және иммунологиялық қасиетін анықтайды және штамы вирулентті. Капсуласы жұқа болған сайын вируленттігі жоғары және бактерия патогенді. Кейбір патогенді бактериялар капсуласыз авирулентті болып келеді.
Кейбір жағдайда клетка сыртындағы гликандарға көміртегі көзі мен энергия қызметін атқарады, ал азот құрайтын полисахаридтер продуцент үшін азот көзі болады. Полианионды полисахаридтер қоршаған ортадан катионмен концентрлейді және клеткаға оны тасымалдауға қабілетті. Н-алканды ортада өскен микроорганизмнің полисахаридті – липидті комплексі: псевдомонад, нокардий, коринебактерия, ашытқының жақсы эмульгаторы. Олар беткі жағының керілгенін төмендетеді, мицеллияның пайда болуына, клеткаға көмірсудың енуіне мүмкіндік жасайды.
Бейтарап клетка сыртындағы полисахаридтер клетканың жиналуын және жіпше тәрізді формасының бүтін болуын қолдайды, Полисахаридтер белгілі заряды бар клетканың диспергирленуіне қарсы бола алады. Кейбір бактериялар экзогликандар клетканың бетіне жабысуына жауапты. Демек, Str, mutans, тісте кариестің болуын тудырады, декстран арқылы тістің эмалына жабысады.
Полисахаридтердің биосинтезі
Гликандар биосинтезі моносахаридтердің арасында глюкозидтті байланыстың түзілуіне әкеліп соғады. Жалпы түрде бұны былай түсінуге болады, глюкозильді донор глюколизді акцептор – затравкаға береді де, ал өзі босайды(сурет).
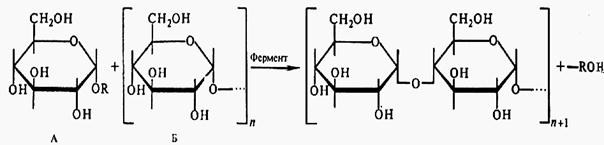
А – донор; Б – акцептор
62 сурет - Глюкозидті байланыстың түзілуінің сызба – нұсқасы
Полимеризация дайын полисахаридтер түзілгенге дейін жүреді. Процесс арнайы гликозилтрансферазалармен катализденеді, ал полимердің басқа толық түзілмеген полисахаридтің сызықша тізбегінің фрагментін ыдыратып, оны дәл сол немесе соған ұқсас (аналогы) тізбегінің белгілі бір жеріне ауыстырып, басқа гликозилтрансферазалармен катализденеді (сурет).
Гетерополисахаридтер биосинтезі жағдайынан екі мүмкіндік туады: әртүрлі мономер бірлігінің кезектесіп реттеліп келуі немесе донорға байланысып қалатын ди-және олигосахаридтердің алдын – ала синтезделуі кейіннен полимеризациялануы.
Полисахаридтер биосинтезінде акцептор болып олигосахар және толығымен құрастырылмаған гликандар келіп түседі. Көбінесе біріншілік акцептор болып олигосахар саналады. Бұл мысалға, декстрандар мен ливандардың (сахароза), целлюлозалардың (целлодекстриндерге), хитиннің (хитодекстриндер) синтезінде болады. Кейде біріншілік акцептор тек толығымен құрастырылмаған полисахарид – «затравка» ғана бола алады.
<







