 2015-04-30
2015-04-30 504
504После образования инициаторного комплекса активируется следующий кодон матричной РНК. Он спаривается с антикодоном транспортной РНК, несущей соответствующую аминокислоту, закодированную в триплете.
Первая аминокислота в полипептидной цепи, метионин, соединяется со следующей с помощью пептидной связи и отщепляется от транспортной РНК.
Свободная транспортная РНК покидает рибосому. Рибосомный комплекс перемещается вдоль молекулы матричной РНК, и комплекс транспортной РНК с присоединенными аминокислотами занимает освобожденное место.
Следующий конец матричной РНК становится активированным и спаривается с соответствующим антикодоном транспортной Р НК, несущим следующую аминокислоту. Эти события повторяются до тех пор, пока рибосома не дойдет до конца (рис. 8.16).
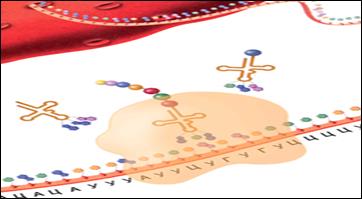
Рис. 8.16. Конечная точка прохождения рибосомы
Введение генов в бактерию и их экспрессия.
Для введения генов в бактерии фрагменты геномной ДНК изменяют, удаляя из них некодирующие области и участки соседних генов, в следующей последовательности:
1. Для введения генов в клетки прокариот подбирают соответствующий вектор, способный проникнуть в клетку реципиента. Чаще всего используют ДНК–фаг или плазмиду бактерий.
2. Плазмиду разрезают рестриктазо й для образования «липких концов».
3. При необходимости производят надстройку концов ДНК у вектора, чтобы они были комплементарны концам ДНК вводимого гена.
4. ДНК донора разрушают рестриктазами для выделения генов.
5. Для введения генов в бактерии фрагменты геномной ДНК изменяют, удаляя из них некодирующие области и участки соседних генов.
6. При объединении в одном растворе ДНК вводимого гена и ДНК вектора, комплементарные концы «слипаются» и образуются водородные связи. С помощью ДНК-лигазы сшивают цепи гибридной ДНК.
7. Образованная гибридная молекула вводится в клетку бактерии-реципиента.
8. У прокариот структурный ген представляет собой непрерывный участок молекулы ДНК, а включение и выключение транскрипции обеспечивается дополнительными факторами транскрипции, которые связываются с соответствующими участками ДНК.
В бактериальной клетке содержится 2-3 тысячи различных белков, каждая из форм которых составляет в среднем 0,04% от их общей массы. При клонировании и экспрессии в бактериях чужеродного гена доля кодируемого им индивидуального белка может достигать 40%. Клетки, в которых эта величина составляет 2% и более, называется суперпродуцентами. В целях создания оптимальных условий для наращивания культуры и синтеза в ней высоких концентраций белков, экспрессию клонируемых генов у бактерий продуцентов включают только на поздних стадиях их развития.
Экспрессия генов в дрожжах.
Дрожжевые клетки удобно использовать для получения разнообразных продуктов промышленного назначения. Преимуществами дрожжей, по сравнению с бактериями, является их рост на дешёвых субстратах и высокая генетическая стабильность при промышленном культивировании. Экспериментировать с дрожжевыми клетками легко, так как они имеют сравнительно малый размер генома и короткое время регенерации (несколько часов). В качестве объекта для клонирования и экспрессии рекомбинантных ДНК чаще всего используют Saccharomyces cerevisiae (рис. 8.17; 1 –экспрессирование; 2 - клонирование).
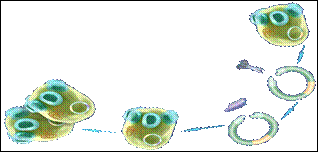
Рис. 8.17. Схема применения Saccharomyces cerevisiae








