 2015-04-23
2015-04-23 3537
3537Стійкий стан (англ. steady - state). При виконанні вправ постійної аеробної потужності за періодом швидких змін функцій організму (впрацювання) настає період, стійкого стану. При виконанні вправ невеликої потужності впродовж періоду стійкого стану є кількісна відповідність між потребою організму в кисні (кисневим запитом) і її задоволенням. Тому такі вправи є вправами з істинно стійким станом. Кисневий борг після нетривалого їх виконання практично дорівнює лише кисневому дефіциту, що виникає спочатку роботи.
У всіх вправах аеробної потужності з рівнем споживання О2 більше 50% від МСК, як і у всіх вправах анаеробної потужності, не можна виділити робочий період з істинно стійким, незмінним станом функцій ні по споживанню О2, ні тим більше за іншими показниками. Це пов'язано з тим, що протягом всього часу їх виконання, не дивлячись на швидке підвищення ступеня споживання О2, потужність системи аеробного окислення відстає від фізичної потужності і не в змозі досягти показників, необхідних для повного покриття кисневого боргу. Таким чином, починаючи з аеробних вправ великої потужності вже немає повної рівноваги між кисневим запитом і його задоволенням під час самої роботи. Тому після них реєструється кисневий борг, який тим більше, чим більша потужність роботи і її тривалість. Але до певного моменту наростаючий кисневий борг не призводить до зниження демонстрованої спортсменом фізичної потужності вправи. Це явище характеризується як псевдо (квазі) стійкий стан або період з повільними функціональними змінами ("дрейфом").
В період квазістійкого стану організму відбувається поступова компенсаторна перебудова в діяльності серцево-судинною, дихальної, нервово-м'язової, ендокринної і інших систем (рис. 3), яка дозволяє підтримувати задані параметри потужності навантаження.
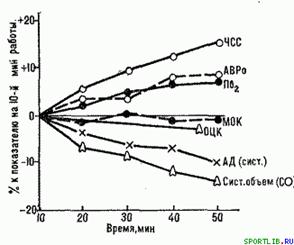
Рис. 3. Зміна ("дрейф") показників діяльності серцево-судинної системи впродовж субмаксимальної аеробної роботи. За 0 прийняті показники на 10-ій хв. роботи.
4. Процес стомлення
Процес стомлення - це сукупність змін, що відбуваються в різних органах, системах і організмі в цілому, в період виконання фізичної роботи що приводить врешті-решт до неможливості її продовження. Стан втоми характеризується викликаним роботою тимчасовим зниженням працездатності, яке виявляється в суб'єктивному відчутті втоми. В стані втоми людина не здатна підтримувати необхідний рівень інтенсивності і (або) якості (техніка виконання) роботи або вимушена відмовитися від її продовження.
Ступінь участі тих або інших фізіологічних систем у виконанні вправ різного характеру і потужності неоднаковий. У виконанні будь-якої вправи можна виділити основні, ведучі, найбільш завантажувані системи, функціональні можливості яких визначають здатність людини виконати дану вправу на необхідному рівні інтенсивності і (або) якості. Ступінь завантаженості цих систем по відношенню до їх максимальних можливостей визначає граничну тривалість виконання даної вправи, тобто період настання стану втоми. Таким чином, функціональні можливості провідних систем не тільки визначають, але і лімітують інтенсивність і граничну тривалість і (або) якість виконання даної вправи.
При виконанні різних вправ причини втоми неоднакові. Розгляд основних причин втоми пов'язаний з двома основними поняттями. Перше поняття - локалізація втоми, тобто виділення тієї провідної системи (або систем), функціональні зміни в якій і визначають настання стану втоми. Друге поняття - механізми втоми, тобто ті конкретні зміни в діяльності провідних функціональних систем, які обумовлюють розвиток втоми.
По локалізації втоми можна, по суті, розглядати три основні групи систем, що забезпечують виконання будь-якої вправи:
- регулюючі системи - центральна нервова система (при потужних навантаженнях), вегетативна нервова система і гормонально-гуморальна система (при роботі на витривалість);
- система вегетативного забезпечення м'язової діяльності - системи дихання, крові і кровообігу (роль, особливо велика при тривалих вправах);
- виконавча система - руховий (периферичний нервово-м'язовий) апарат (виснаження енергетичних ресурсів, отруєння продуктами розпаду енергетичних речовин, що накопичуються в результаті недостатнього надходження кисню).
Для різних вправ характерна специфічна комбінація провідних систем (локалізації) і механізмів стомлення.
При виконанні вправ максимальної анаеробної потужності (до 10 с.) найбільш важливу роль в розвитку втоми грають процеси, що відбуваються в ЦНС і виконавчому нервово-м'язовому апараті.
Під час цих вправ вищі моторні центри повинні активувати максимально можливе число спинальних мотонейронів працюючих м'язів і забезпечити високочастотну імпульсацію. Така інтенсивна "моторна команда" може підтримуватися лише протягом декількох секунд. Особливо рано знижується частота імпульсації, і відбувається виключення швидких мотонейронів. Виключно швидко витрачаються фосфагени в працюючих м'язах, особливо креатинофосфат, так що одним з провідних механізмів стомлення при виконанні цих вправ служить виснаження фосфагенів як основних субстратів, здатних забезпечувати таку роботу. Анаеробний гліколіз розвивається повільніше, тому за декілька секунд роботи концентрація лактату в м'язах, що скорочуються, збільшується незначно. Системи вегетативного забезпечення зважаючи на їх інертність не грають вирішальної ролі у виконанні цих вправ і відповідно в розвитку втоми.
При виконанні вправ майжемаксимальной анаеробної потужності ( тривалість 20-50 с. ) розвиток втоми також визначають зміни що відбуваються в ЦНС і у виконавчому м'язовому апараті. Як і при максимальній анаеробній роботі, ЦНС повинна забезпечувати рекрутування і високочастотну імпульсацію більшості спинальних мотонейронів, що іннервують основні робочі м'язи. У самих м'язових клітинах відбувається інтенсивне витрачання субстратів анаеробного метаболізму - фосфагенів і м'язового глікогену, накопичується і діффундірує в кров значна кількість молочної кислоти. Отже разом з виснаженням фосфагенов важливою причиною стомлення при майжемаксимальній анаеробній роботі є накопичення в м'язах і крові молочної кислоти, що, з одного боку, знижує швидкість глікогеноліза в м'язах, а з іншої - несприятливо впливає на діяльність ЦНС.
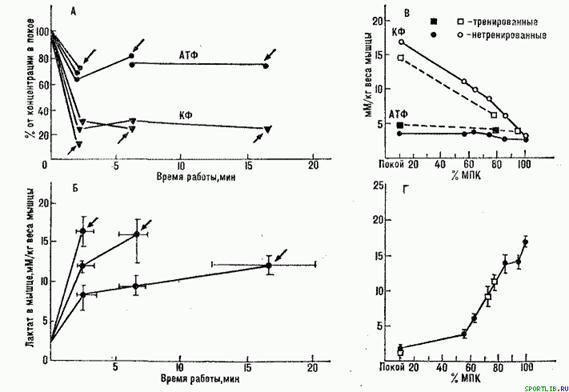
Рис. 4. Концентрація фосфагенів (Л) і лактату (Б) в робочих м'язах впродовж анаеробних і аеробних вправ різної граничної тривалості (стрілки відповідають моменту відмови від роботи); концентрація фосфагенів (В) і лактату (Г) в робочих м'язах після вправ різної відносної потужності аероба (по. Д. Карлссону та ін., 1971)
Під час виконання вправ субмаксимальної анаеробної потужності (тривалість 1-2 хв.)ресинтез фосфагенів відбувається з достатньою швидкістю, тому в кінці роботи не виявляється помітного їх витрачання (рис. 4 А). Головним механізмом стомлення в цих вправах служать накопичення лактату в м'язах (рис. 4 Б) та крові і обумовлене ним зниження рН в м'язових клітинах і крові. Обидва ці чинника приводять до зменшення швидкості глікогеноліза в м'язах і негативно впливають на діяльність ЦНС. При роботі субмаксимальної анаеробної потужності додатковим (хоча не дуже істотним) чинником, лімітуючим працездатність, служать функціональні можливості, кисневотранспортної системи. Тому одним з механізмів стомлення при виконанні такої роботи є недостатнє постачання кисню м'язам.
При виконанні вправ максимальної аеробної потужності (тривалість 3-10 хв. ) втома пов'язана перш за все з кисневодтранспортною системою, граничні можливості якої є чинником, лімітуючим працездатність. Один з головних механізмів стомлення в даному випадку - недостатнє забезпечення працюючих м'язів киснем. В процесі такої роботи значну частину енергії м'язи отримують в результаті анаеробного глікогеноліза з утворенням молочної кислоти, накопичення якої (зниження рН) в м'язах і крові також грає важливу роль в розвитку втоми.
Виконання вправ майжемаксимальної аеробної потужності (тривалістьдо 30 хв. ) також лімітується в основному можливостями кисневотранспортної системи. В процесі їх виконання концентрація фосфагенів знижується незначно, концентрація лактату в м'язах і крові відносно невелика (рис. 4). Втома пов'язана із зниженням продуктивності серцево-судинної системи, особливо серця. Серцева продуктивність виступає як головний чинник, лімітуючий постачання м'язам кисню. Робота забезпечується переважно глікогенолізом. Проте відмова від продовження її прямо не пов'язана з виснаженням вуглеводних ресурсів організму (рис. 5). Висока концентрація молочної кислоти в м'язах і крові дозволяє розглядати її як один з важливих механізмів стомлення при виконанні вправ майжемаксимальної аеробної потужності.
Вправи субмаксимальної аеробної потужності (тривалість до 120 хв. ) пов'язані з великим навантаженням на серцево-судинну систему. Їх виконання забезпечуєтся окислювальними процесами в працюючих м'язах, що використовують як основний субстрат м'язовий глікоген і глюкозу крові. Головним механізмом стомлення при таких вправах служить виснаження запасів глікогену в працюючих м'язах і печінці (рис. 5). Більшість змін, які спостерігаються в діяльності серцево-судинної системи, впродовж періоду квазістійкого стану (рис. 2), відображають перебіг процесів, які, врешті-решт, приводять до втоми. Велике і тривале навантаження на серце веде до зниження продуктивності міокарду. Певну роль в розвитку втоми грають вимоги, що підвищуються по мірі продовження роботи, до підтримки необхідної температури тіла (робочої гіпертермії).
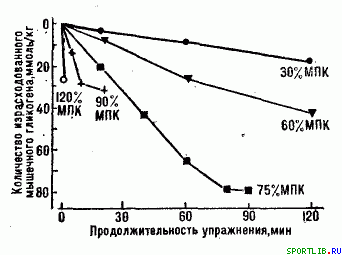
Рис. 5. Зміст м'язового глікогену після вправ різної відносної потужності і (відповідно) різної граничної тривалості. Числа біля кривих указують, відносну потужність роботи у відсотках від МСК - 120% МСК відповідає, майжемаксимальной анаеробної потужності
Вправи середньої аеробної потужності (тривалістьдо декількох годин ) також дають велике навантаження на кисневотранспортну систему. При роботі такої потужності відбувається значна витрата глікогену м'язів і посилена витрата (виснаження) глікогену печінки, що веде до розвитку гіпоглікемії. Таким чином, повторно страждає ЦНС, для якої глюкоза крові грає роль єдиного енергетичного джерела. Крім того, велике значення має порушення процесів терморегуляції, що може викликати критичне підвищення температури тіла. В результаті додаткового перерозподілу кровотоку (посилення шкірного кровотоку і зниження кровотоку працюючих м'язів) відбувається підвищення тепловіддачі. Доставка кисню до робочих м'язів знижується, що веде до м'язової втоми.
Вправа малої аеробної потужності ( впродовж декількох годин при потужності роботи менше 50% від індивідуального МСК ) значною мірою характеризуються тією ж локалізацією і механізмами стомлення, що і вправи середньої аеробної потужності. Відмінність полягає в повільнішому настанні описаних процесів і в більшому витрачанні жирів, недоокислені продукти розщеплювання яких можуть поступати в кров і бути важливим чинником стомлення.
6. Відновлення
Після припинення вправи відбуваються зворотні зміни в діяльності тих функціональних систем, які забезпечували виконання даної вправи. Вся сукупність змін в цей період об'єднується поняттям відновлення. Впродовж відновлювального періоду видаляються продукти робочого метаболізму і поповнюються енергетичні запаси, пластичні (структурні) речовини (білки та ін.) і ферменти, витрачені за час м'язової діяльності. По суті, відбувається відновлення порушеного роботою гомеостазу. Проте відновлення - це не тільки процес повернення організму до передробочого стану. У цей період відбуваються також зміни, які забезпечують підвищення функціональних можливостей організму, тобто позитивний тренувальний ефект.
У періоді відновлення можна виділити 4 фази: 1) швидкого відновлення, 2) повільного відновлення, 3) суперкомпенсації (або "надвідновлення"), 4) тривалого (пізнього) відновлення. Наявність цих фаз, їх тривалість і характер сильно варіюють для різних функцій. Першим двом фазам відповідає період відновлення працездатності, пониженої в результаті важкої роботи, третій фазі - підвищена працездатність, четвертій - повернення до нормального (передробочого) рівня працездатності (рис. 6).
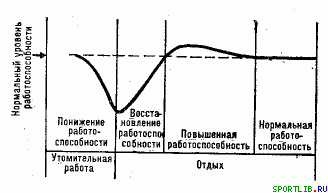
Рис. 6. Схематичне представлення зміни працездатності впродовж роботи до відмови і в період відновлення (по Р. Ст. Фольбортту)
Загальні закономірності відновлення функцій після роботи полягають в наступному.
- По-перше, швидкість і тривалість відновлення більшості функціональних показників знаходяться в прямій залежності від потужності роботи: чим вище потужність роботи, тим більші зміни відбуваються за час роботи і (відповідно) тим вище швидкість відновлення.
- По-друге, відновлення різних функцій протікає з різною швидкістю, а деякі фази відновного процесу і з різною спрямованістю, так що досягнення ними рівня спокою відбувається неодночасно (гетерохронно).
- По-третє, працездатність і багато функцій організму, визначаючих її впродовж періоду відновлення після інтенсивної роботи не тільки досягають передробочого рівня, але можуть і перевищувати його, проходячи через фазу "надвідновлення " ( рис. 6). Коли мова йде про енергетичні субстрати, то таке тимчасове перевищення передробочого рівня носить назву суперкомпенсації (Н. Н. Яковлєв).
В процесі м'язової роботи витрачаються кисневий запас організму, фосфагени (АТФ і КРФ), вуглеводи, (глікоген м'язів і печінки, глюкоза крові) і жири. Після роботи відбувається їх відновлення. Виняток становлять жири, відновлення яких може і не бути.
У сукупному кисневому боргу виділяють швидкий і повільний компоненти.
Швидкий (алактатный) компонент О2-боргу пов'язаний головним чином з використанням О2 на швидке відновлення витрачених за час роботи високоенергетичних фосфагенів в робочих м'язах (впродовж 30 с після припинення роботи відновлюється до 70%), а також з відновленням нормального вмісту О2 у венозній крові (до 0,2 л, на перших секундах) і з насиченням міоглобіну киснем (за декілька секунд).
Повільний (лактатный) компонент О2-борга пов'язаний з багатьма чинниками. Великою мірою він пов'язаний з післяробочим усуненням лактату з крові і тканинних рідин. Кисень в цьому випадку використовується в окислювальних реакціях, що забезпечують ресинтез глікогену з лактату крові (головним чином, в печінці і частково в нирках) і окислення лактату в серцевій і скелетних м'язах. Крім того, тривале підвищення споживання О2 пов'язане з необхідністю підтримувати посилену діяльність дихальної і серцево-судинної систем в період відновлення, посилений обмін речовин і інші процеси, які обумовлені підвищеною активністю симпатичної, нервової і гормональної систем, підвищеною температурою тіла, що поволі знижуються впродовж періоду відновлення.
Відновлення глікогену, пластичних (структурних) речовин (білків та ін.) і ферментів, витрачених за час м'язової діяльності, займає більш тривалий час і багато в чому визначається ступенем їх витрачання і характером харчового раціону. Наприклад, після дуже значного (більше 3/4 початкового складу), аж до повного, виснаження глікогену в робочих м'язах його відновлення в перші години при звичайному харчуванніі йде дуже повільно, і для досягнення передробочого рівня потребує до 2 діб. При харчовому раціоні з високим вмістом вуглеводів (більше 70% добового калоражу) цей процес прискорюється - вже за перші 10 год. в робочих м'язах відновлюється більше половини глікогену, до кінця доби відбувається його повне відновлення, а в печінці зміст глікогену значно перевищує звичайне. Надалі кількість глікогену в робочих м'язах та печінці продовжує збільшуватися і через 2-3 дні після "виснажливого" навантаження може перевищити передробочу в 1,5-3 рази - явище суперкомпенсації.
При відновленні білкових структур спостерігається така ж тенденція, що підтверджується набагато більшим вмістом білка в харчовому раціоні представників силових видів спорту (у бодібілдерів кількість споживаного білка досягає 2 г на кілограм ваги спортсмена).

