 2015-04-30
2015-04-30 4214
4214Углеводы подразделяются на три основные группы: моносахариды, олигосахариды и полисахариды.
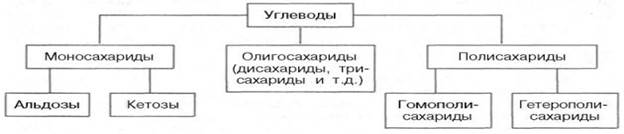
Моносахариды — органические соединения, одна из основных групп углеводов; самая простая форма сахара; являются обычно бесцветными, растворимыми в воде, прозрачными твердыми веществами. Некоторые моносахариды обладают сладким вкусом. Моносахариды — стандартные блоки, из которых синтезируются дисахариды, (такие, как сахароза, мальтоза, лактоза) олигосахариды и полисахариды (такие, как целлюлоза и крахмал), содержат гидроксильные группы и альдегидную (альдозы) или кетогруппу (кетозы). Каждый углеродный атом, с которым соединена гидроксильная группа (за исключением первого и последнего) является хиральным, давая начало многим изомерным формам. Например, галактоза и глюкоза — альдогексозы, но имеют различные химические и физические свойства. Моносахариды представляют собой производные многоатомных спиртов, содержащие карбонильную группу — альдегидную или кетонную.В зависимости от числа атомов углерода подразделяются на триозы, тетрозы, пентозы, гексозы. Для человека наиболее важны глюкоза, фруктоза, галактоза, рибоза, дезоксирибоза.
Продуктами окисления моносахаридов, наряду с альдоновыми и сахарными кислотами, являются уроновые кислоты.
34.Механизмы активации моносахаридов-образование фосфорных эфир нуклеозиддифосфатсахаров (НДФС). Взаимопревращения фосфорных эфиров моносахаридов. Образование пентоз у растений.
Одна из наиболее важных реакций фосфорной кислоты — это присоединение ее к сахарам с образованием сложных эфиров. Самый распространенный фосфорный эфир — глюкоза-6-фосфат — синтезируется в растениях путем переноса фосфорной кислоты с аденозинтрифосфата (АТФ) на глюкозу при одновременном образовании аденозиндифосфата (АДФ). Пентоза — общее родовое химическое название класса пятиуглеродных сахаров, то есть сахаров, общей формулой которых является C5(H2O)5, или C5H10O5. Широко распространены в природе, встречаются в свободном виде, входят в состав гликозидов, полисахаридов. Являются структурными компонентами нуклеиновых кислот.
35.Гликозиды и дубильные вещества, их роль в пищевой промышленности. Специфичность действия гликозидаз. Глюкозоизомераза и современные технологии производства сахаристых веществ. Гликозиды — большая группа летучих безазотистых веществ, состоящих из соединений глюкозы с различными органическими веществами. От гликозидов зависит вкус и аромат некоторых растительных продуктов. В отличие от алкалоидов гликозиды могут быстро разрушаться при хранении продукта ферментами самих растений (ауто-ферментация), а также под действием различных физических факторов, например, при кипячении.При кипячении с водой или под влиянием ферментов гликозиды распадаются на сахаристую и несахаристую часть, называемую агликоном и имеющую различное химическое строение. Именно агликон определяет физиологическую активность гликозидов и ее характер. В практической медицине обычно используются следующие группы гликозидов: сердечные гликозиды, антра-гликозиды, сапонины, горечи, флавоноидные гликозиды и др. Наиболее важное значение имеют сердечные гликозиды. Сердечные гликозиды содержатся в таких растениях, как различные виды наперстянки, ландыш, горицвет и др. Однако, при этом надо иметь в виду, что растения, содержащие сердечные гликозиды, как правило, являются высокотоксичными, ядовитыми. Лекарственное применение находят и некоторые другие гликозиды. Так, в листьях груши, брусники, толокнянки содержится гликозид арбутин, обладающий антибактериальными свойствами. В связи с этим его используют при лечении воспалительных заболеваний мочеполовых путей.Довольно широкое применение в медицине получили так называемые антрагликозиды, содержащиеся в крушине, ревене, кассии, алоэ и других растениях. Эти гликозиды оказывают слабительное действие. Они малотоксичны, стойки при хранении, большинство из них окрашено в красно-оранжевый цвет. Некоторые растения содержат так называемые горькие гликозиды, используемые в медицине как горечи для повышения аппетита и улучшения пищеварения, так как они стимулируют выделение пищеварительных соков. Горькие гликозиды присутствуют в полыни, одуванчике, горечавке, золототысячнике и др. Довольно большую группу гликозидов составляют сапонины, которые содержатся во многих растениях, особенно относящихся к семействам гвоздичных и первоцветных. При взбалтывании с водой сапонины образуют довольно стойкую пену. Сапониносодержащие растения используются в медицине как отхаркивающие (корни истода, синюхи и первоцвета), мочегонные (трава почечного чая), желчегонные (трава зверобоя).Некоторые сапонины обладают свойством понижать артериальное давление, вызывать рвоту, оказывать потогонное действие и т.д. В последние годы выявлено противосклеротическое действие некоторых сапонинов.В последнее время большое значение приобрела группа так называемых флавоноидных гликозидов. Название этих веществ указывает на их желтую окраску, что и послужило основой для их названия («флавум» — по латыни означает желтый). Желтые и оранжевые цвета многих растений зависят от содержащихся в них флавоновых гликозидов, или их агликонов, которые являются производными флавона или оксифлавонов. Например, в табаке, луковой шелухе, пыльце кукурузы, листьях чая, хмеле и некоторых других растениях окраска зависит от кверцетина, агликона гликозида кверцитина. Из флавоноидов создаются лечебные и питательные препараты. В некоторых растениях в чистом виде или в соединениях с сахарами в виде гликозидов содержатся кумарины и фурокумарины. В воде эти соединения обычно плохо растворимы, чувствительны к свету. Чаше кумарины содержатся в растениях семейств зонтичных, бобовых, рутовых, причем концентрируются преимущественно в плодах и корнях. К настоящему времени выделено и изучено более 150 кумаринопроизводных соединений; наиболее важные из них относятся к фурокумаринам. Некоторые из них используются как сосудорасширяющиеся и спазмолитические, другие — как эстрогены, противоопухолевые и фотосенсибилизирующие средства. Очень близко к флавоновым гликозидам стоят гликоиды-антоцианы. Весьма распространен агликон цианидин, входящий в состав красящих веществ плодов вишни, сливы, черной смородины, брусники, некоторые антоциановые производные применяются как слабительные средства, а также по другим показаниям. Дубильные веществаДубильные вещества — соединения различной химической природы. Для них характерна особенность видоизменять коллоидное состояние белков: осаждать их из растворов, вызывать затвердение, противодействовать набуханию в воде и т.д. Различают органические и минеральные дубильные вещества. Органические дубильные вещества бывают животного и растительного происхождения; могут получаться и искусственным путем. Дубильные вещества органического происхождения — это в основном высоконепредельные жиры морских животных, растительного происхождения — это наиболее обширная группа дубителей. Добываются они из дубильных растений.Содержание дубильных веществ в частях одного и того же растения неодинаково: особенно большое количество их бывает в коре и корнях. Содержатся они главным образом в высших растениях. Их роль в физиологии растений полностью еще не выяснена. В медицине дубильные вещества употребляются как противоядие при отравлении алкалоидами, желудочно-кишечных заболеваниях, при ожогах (таннин) и т.д. Большое значение имеют дубильные вещества в пищевой промышленности, где они нередко определяют характер переработки и вкусовые достоинства готового продукта. Например, производство черного чая основано на превращении дубильных веществ, содержащихся в молодых побегах чайного растения в количестве 18—30% (на сухой вес). От правильно проведенного окисления дубильных веществ при ферментации зависит в основном цвет, вкус и аромат чая. Дубильные вещества используются также в производстве какао и ряда других пищевых продуктов. Специфичность действия гликозида зависит от агликона (безуглеводная часть молекулы), но углеводная часть молекулы (гликон) существенно влияет на растворимость, усвоение гликозида организмом и, в конечном счете, проявление и выраженность биологического действия. Важное значение имеет гликозидная связь между двумя частями молекулы, которая включает в себя принадлежащие агликону различные атомы - кислорода, углерода, серы или азота и имеет различную изомерную (альфа- или бета-) конфигурацию. Расщепление связи и, соответственно, утрата активности гликозида связана с большой группой ферментов - гликозидаз, определяемых в зависимости от природы Сахаров (глюкозидазы, фруктозидазы и др.) и их и зомерных характеристик.
Глюкозоизомераза (ГИ) — индуцибельный фермент. У большинства микроорганизмов он накапливается внутри клеток Но имеются культуры, продуцирующие внутри- и внеклеточные изомеразы. Для некоторых из них, доказана идентичность синтезируемых ими эндо- и экзоферментов. Для внутриклеточных глюкозоизомераз важно выявление локализации этих ферментов внутри клеток. В области клеточной стенки и периплазматического пространства обычно расположены ферменты, выполняющие функцию превращения составных частей окружающей среды в необходимые для метаболизма соединения. Очевидно, что можно отнести и к ГИ, функционирование которой в клетках (совместно с пермеазами и киназами соответствующих сахаров) приводит к образованию ключевых продуктов метаболизма, таких, как D-глюкозо-6-фосфат и D-ксилоза-5-фосфат. Наибольший выход ГИ из клеток достигается при обработке их ультразвуком, разрушении биомассы под давлением или автолизе в присутствии различных добавок (детергенты, толуол, лизоцим,. стрептолитин). Представляет интерес возможность выделения ГИ из клеток действием на них иммобилизованными литическими ферментами микробного происхождения. Выделенную из биомассы глюкозоизомеразу (ГИ) очищают или иммобилизуют. При частичной очистке используют тепловую или кислотную денапрации. фракционное осаждение органическими растворителями, сульфатом аммония, а также солями Мg2+ или Мn2+. Большинство исследователей для частичной очистки ГИ используют комбинации перечисленных методов. Как и у других термостабильных ферментов в молекуле большинства ГИ присутствуют преимущественно остатки аргинина, глицина и группы, образующие а-спираль (аланин, лейцин, глутаминовая кислота и аспаргин). В тоже время наблюдается пониженное содержание остатков цистеина, серина и лизина. Особенность многих термостабильных внутриклеточных ферментов - пониженое содержание или полное отсутствие цистеина. Это объясняется восстановительной способностью внутриклеточной среды, сводящей к минимуму окислительные процессы. Для многих ферментов с низким содержанием цистеина, в том числе для ГИ, кофакторы - мультивалентные ионы. Необходимо отметить, что ГИ - это исключение из общей закономерности, согласно которой между термоустойчивостью ферментов и температурой выращивания продуцирующих их микроорганизмов существует строгая корреляция. Как и для ряда термостабильных ферментов мезофильных грибов, для ГИ этот факт можно объяснить существованием нескольких конформационных состояний. Одному или некоторым из них при повышенной температуре свойственна большая стабильность. Происходящие при этом структурные перестройки не затрагивают экранируемый активный центр изомеразы и его микроокружение, благодаря чему фермент обладает способностью катализировать реакции изомеризации при повышенной температуре. ГИ обладает групповой специфичностью. Фермент может изомеризовать в основном два субстрата: D -глюкозу и D -ксилозу; в меньшей степени— D -рибозу, а также D -арабинозу, D -аллозу, D -рамнозу и L -арабинозу. Наибольшую степень сродства ГИ проявляет к D-ксилозе, хотя известны случаи, когда изомераза за обнаруживает почти одинаковое сродство как к D-ксилозе, так и к D-глюкозе.Разработаны принципиально новые технологии проведения традиционных процессов получения сахаристых веществ для пищевой отрасли — инвертного сахара и крахмальных паток. Полученные по новым технологиям сахаристые вещества имеют хорошие потребительские качества и отличаются высокой чистотой и отсутствием побочных веществ. Так, инвертные сахарные сиропы не провоцируют возникновение кариеса, обладают по сравнению с сахаром более мягким вкусом, меньшей вязкостью и большим содержанием сахаристых веществ (можно приготовить 79,5 %-ые сиропы при 20оС). Крахмальные патоки, полученные по новым технологиям, имеют заданный, легко контролируемый в процессе гидролиза состав, который удовлетворяет требованиям любого потребителя (кондитеров, пивоваров, специалистов детского питания и др.). Существующие в перерабатывающей промышленности гомогенные технологии, в основном, с использованием неорганических кислот, заменены на гетерогенные технологии с участием ферментов — природных биокатализаторов. Модернизирована стадия подготовки крахмал-содержащего сырья к последующему гидролизу. При “сухой” декстринизации крахмала существенно упрощается процесс и уменьшается количество стадий по сравнению с традиционной технологией. При получении декстринизированного пшеничного крахмала используется мука из некондиционного зерна, запасы которого в России поистине огромны, а отделенная клейковина может быть использована в хлебопечении для улучшения качества муки. Получаемые по гетерогенным технологиям конечные продукты переработки сырья отличаются сравнительно высокой чистотой и отсутствием побочных веществ. Возможен строгий контроль за глубиной гидролиза крахмала и составом получаемых паток по требованию потребителя.
36.Олигосахариды. Важнейшие представители дисахаридов(сахароза, мальтоза, целлобиоза, лактоза) и трисахаридов (рафиноза, мальтотриоза, паноза). Их характеристика. ОЛИГОСАХАРИДЫ - углеводы, молекулы к-рых построены из неск. моносахаридных остатков (от 2 до 10-20), соединенных гликозидными связями (экзоциклич. связь ано-мерного атома С моносахарида с атомом О соседнего моносахаридного остатка). В соответствии со степенью полимеризации различают дисахариды (биозы), трисахари-ды (триозы), тетрасахариды (тетраозы) и т.д.В состав О. могут входить остатки к.-л. одного моносахарида (гомоолигосахариды) или разных моносахаридов (гете-роолигосахариды). Каждый моносахаридный остаток может находиться в одной из четырех возможных циклич. форм (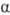 -и
-и 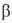 -фураноза,
-фураноза, 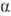 - и
- и 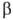 -пираноза)и соединяться гликозидной связью с любой гидроксильной группой соседнего остатка (включая полуацетальный гидроксил). Отсюда следует, что даже из двух одинаковых гексоз можно построить 30, а из двух разных гексоз-56 изомерных диса-харидов; три разные гексозы теоретически дают 4896 изомерных трисахаридов; с ростом степени полимеризации число возможных изомеров быстро достигает астрономич. величин.Если в молекуле О. все гликозидные связи образованы полуацетальным гидроксилом одного и спиртовым гидро-ксилом другого моносахаридного остатка, в конце цепи остается один незамещенный полуацетальный гидроксил, за счет к-рого О. проявляет св-ва карбонильных соед., характерные для моносахаридов (р-ции окисления и восстановления, мутаротацию и др.); такие О. наз. восстанавливающими (редуцирующими). Если же один из моносахаридных остатков в молекуле О. связан гликозидной связью с полуацетальным гидроксилом другого моносахарида, такие О. не содержат полуацетального гидроксила и наз. невосстанавливающими (нередуцирующими). О., в к-рых к спиртовым гидроксилам каждого моносахаридного остатка присоединено не более одного соседнего остатка, наз. линейными (неразветвленными); присоединение двух и более моносахаридов к спиртовым гидроксилам одного и того же моносахаридного остатка приводит к разветвлению О. Т. обр., уже трисахариды могут иметь разветвленную структуру. Моносахаридные остатки, расположенные на концах углеводных цепей, наз. концевыми (терминальными). В восстанавливающих О. различают концевой восстанавливающий моносахарид (он м. б. только один) и концевые невосстанавливающие моносахариды (их на единицу больше, чем точек разветвлений).Строгая номенклатура О. весьма громоздка. Название О. образуется по типу О-замещенных производных моносаха-ридов, исходя из названия восстанавливающего звена с указанием всех имеющихся заместителей; для невосстанавливающих О. номенклатура аналогична номенклатуре гликози-дов. В названиях линейных О. часто применяется последовательное перечисление моносахаридных остатков с указанием типа связи между ними. Весьма употребительны тривиальные названия О., обычно связанные с источником получения в-ва, и способы сокращенной записи структур, в к-рых моносахаридные остатки обозначаются тремя буквами, абс. конфигурации - буквами D или L, размер цикла - буквами f (фураноза) или р (пираноза), конфигурации гликозидных центров-буквами
-пираноза)и соединяться гликозидной связью с любой гидроксильной группой соседнего остатка (включая полуацетальный гидроксил). Отсюда следует, что даже из двух одинаковых гексоз можно построить 30, а из двух разных гексоз-56 изомерных диса-харидов; три разные гексозы теоретически дают 4896 изомерных трисахаридов; с ростом степени полимеризации число возможных изомеров быстро достигает астрономич. величин.Если в молекуле О. все гликозидные связи образованы полуацетальным гидроксилом одного и спиртовым гидро-ксилом другого моносахаридного остатка, в конце цепи остается один незамещенный полуацетальный гидроксил, за счет к-рого О. проявляет св-ва карбонильных соед., характерные для моносахаридов (р-ции окисления и восстановления, мутаротацию и др.); такие О. наз. восстанавливающими (редуцирующими). Если же один из моносахаридных остатков в молекуле О. связан гликозидной связью с полуацетальным гидроксилом другого моносахарида, такие О. не содержат полуацетального гидроксила и наз. невосстанавливающими (нередуцирующими). О., в к-рых к спиртовым гидроксилам каждого моносахаридного остатка присоединено не более одного соседнего остатка, наз. линейными (неразветвленными); присоединение двух и более моносахаридов к спиртовым гидроксилам одного и того же моносахаридного остатка приводит к разветвлению О. Т. обр., уже трисахариды могут иметь разветвленную структуру. Моносахаридные остатки, расположенные на концах углеводных цепей, наз. концевыми (терминальными). В восстанавливающих О. различают концевой восстанавливающий моносахарид (он м. б. только один) и концевые невосстанавливающие моносахариды (их на единицу больше, чем точек разветвлений).Строгая номенклатура О. весьма громоздка. Название О. образуется по типу О-замещенных производных моносаха-ридов, исходя из названия восстанавливающего звена с указанием всех имеющихся заместителей; для невосстанавливающих О. номенклатура аналогична номенклатуре гликози-дов. В названиях линейных О. часто применяется последовательное перечисление моносахаридных остатков с указанием типа связи между ними. Весьма употребительны тривиальные названия О., обычно связанные с источником получения в-ва, и способы сокращенной записи структур, в к-рых моносахаридные остатки обозначаются тремя буквами, абс. конфигурации - буквами D или L, размер цикла - буквами f (фураноза) или р (пираноза), конфигурации гликозидных центров-буквами 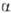 или
или 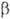 ; цифры в скобках обозначают положения гидроксильных групп, участвующих в межмоно-мерной связи; направление гликозидной связи указывается стрелкой (знак ~ обозначает, что моносахарид может иметь
; цифры в скобках обозначают положения гидроксильных групп, участвующих в межмоно-мерной связи; направление гликозидной связи указывается стрелкой (знак ~ обозначает, что моносахарид может иметь 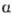 - или
- или 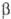 -конфигурацию). Примеры О.: невосстанавливающий дисахарид
-конфигурацию). Примеры О.: невосстанавливающий дисахарид 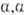 -трегалоза (
-трегалоза (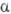 -D-глюкопиранозил-
-D-глюкопиранозил- 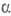 -D-глюкопиранозид; ф-ла Г), восстанавливающие дисахариды лактоза (4-О-
-D-глюкопиранозид; ф-ла Г), восстанавливающие дисахариды лактоза (4-О- 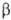 -D-галактопиранозил-D-глюкоза; II) и мальтоза, или солодовый сахар (4-О-
-D-галактопиранозил-D-глюкоза; II) и мальтоза, или солодовый сахар (4-О- 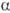 -D-глюкопиранозил-D-глюкоза; III), разветвленный гетеротрисахарид солатриоза (2-О-
-D-глюкопиранозил-D-глюкоза; III), разветвленный гетеротрисахарид солатриоза (2-О- 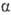 -L-рамнопиранозил-3-О-
-L-рамнопиранозил-3-О- 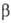 -D-глюкопиранозил-D-га-лактоза; IV), линейный гомотрисахарид мальтотриоза [О-
-D-глюкопиранозил-D-га-лактоза; IV), линейный гомотрисахарид мальтотриоза [О- 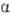 -D-глюкопиранозил-(1
-D-глюкопиранозил-(1 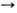 4)-О-
4)-О- 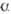 -D-глюкопиранозил-(1
-D-глюкопиранозил-(1 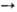
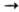 4)-D-глюкоза; V].Главным источником получения разнообразных О. служат р-ции частичного (химического или ферментативного) расщепления прир. полисахаридов, гликолипидов и гликопро-теинов. Большинство О.-бесцв. кристаллич. соед., раств. в воде, меньше раств. в полярных орг. р-рителях и не раств. в неполярных. Невосстанавливающие О. кристаллизуются легко, тогда как восстанавливающие существуют в р-рах в виде смеси таутомерных форм и часто кристаллизуются с большим трудом. Из хим. р-ций О. важнейшей является полный кислотный гидролиз под действием разб. минер. к-т, используемый для определения моносахаридного состава. Восстановление или окисление карбонильной группы позволяет идентифицировать моносахарид. расположенный на восстанавливающем конце молекулы. Превращ. всех гидроксильных групп О. в метиловые эфиры с послед. гидролизом и идентификацией метилир. моносахаридов (т. наз. метод метилирования) служит хим. способом определения размера циклов и расположения межмономерных связей в молекулах О.; для этой же цели используется периодатное окисление. Последовательность моносахаридных остатков и конфигурации отдельных гликозидных центров определяют при расщеплении О. специфич. гликозидазами. Для выделения О. из сложных смесей используют хрома-тографич. методы, причем разделение О. по степени полимеризации достигается с помощью гелъ-хроматографии, а более тонкое разделение изомеров выполняют с применением высокоэффективной жидкостной хроматографии. Низшие О. в виде летучих производных (метиловых, триметилсилило-вых эфиров или ацетатов) удается разделять с помощью гжх.Основным представителем дисахаридов является сахароза. Молекула сахарозы состоит из остатков молекулы D-глюкозы и D-фруктозы. Химическая формула - C12H22O11. Сахароза - один из главных углеводов в организме человека, бесцветное кристаллическое вещество. При температуре выше 200є C разлагается с образованием так называемых карамелей. Сахароза не растворима в неполярных органических растворителях, в абсолютном метаноле и этаноле, умеренно растворима в атилацетате, анилине, в водных растворах метанола и этанола. Хорошо растворима в воде. Сахароза не обладает редуцентными свойствами, поэтому она устойчива к действию щелочей, но гидрализуется под влиянием кислот и ферментов сахараз с образованием D- глюкозы и D- фруктозы. Со щелочным металлами образует сахараты. Сахароза является одним из основных дисахаридов. Она гидролизуется HCl желудочного сока и сахаразой слизистой оболочкой тонкой кишки человека. Сахароза входит в состав сахара (99,75%), используемого для придания пище сладкого вкуса. Сахарозу также называют свекловичным сахаром. Другой представитель дисахаридов - лактоза (молочный сахар). Она состоит из остатков гелактозы и глюкозы. Лактоза - важная составная часть молока млекопитающих и человека. Образуется в процессе лактации в молочной железе из глюкозы и является для новорожденных ее источником. Лактоза облегчает всасывание кальция их кишечника. Содержание лактозы в женском молоке - 7 г/ 100 мл. В молоке коров и коз - 4,5г/100 мл. Мальтоза (солодовый сахар) — промежуточный продукт расщепления крахмала и гликогена в желудочно-кишечном тракте, происходящего под влиянием амилазы — фермента, выделяемого поджелудочной железой. Образующаяся мальтоза расщепляется затем мальтазой кишечного сока до двух остатков глюкозы. В свободном виде в пищевых продуктах мальтоза встречается в меде, солоде, пиве, патоке (мальтозной) и продуктах, изготовляемых с добавлением патоки (хлебобулочные, кондитерские изделия).
4)-D-глюкоза; V].Главным источником получения разнообразных О. служат р-ции частичного (химического или ферментативного) расщепления прир. полисахаридов, гликолипидов и гликопро-теинов. Большинство О.-бесцв. кристаллич. соед., раств. в воде, меньше раств. в полярных орг. р-рителях и не раств. в неполярных. Невосстанавливающие О. кристаллизуются легко, тогда как восстанавливающие существуют в р-рах в виде смеси таутомерных форм и часто кристаллизуются с большим трудом. Из хим. р-ций О. важнейшей является полный кислотный гидролиз под действием разб. минер. к-т, используемый для определения моносахаридного состава. Восстановление или окисление карбонильной группы позволяет идентифицировать моносахарид. расположенный на восстанавливающем конце молекулы. Превращ. всех гидроксильных групп О. в метиловые эфиры с послед. гидролизом и идентификацией метилир. моносахаридов (т. наз. метод метилирования) служит хим. способом определения размера циклов и расположения межмономерных связей в молекулах О.; для этой же цели используется периодатное окисление. Последовательность моносахаридных остатков и конфигурации отдельных гликозидных центров определяют при расщеплении О. специфич. гликозидазами. Для выделения О. из сложных смесей используют хрома-тографич. методы, причем разделение О. по степени полимеризации достигается с помощью гелъ-хроматографии, а более тонкое разделение изомеров выполняют с применением высокоэффективной жидкостной хроматографии. Низшие О. в виде летучих производных (метиловых, триметилсилило-вых эфиров или ацетатов) удается разделять с помощью гжх.Основным представителем дисахаридов является сахароза. Молекула сахарозы состоит из остатков молекулы D-глюкозы и D-фруктозы. Химическая формула - C12H22O11. Сахароза - один из главных углеводов в организме человека, бесцветное кристаллическое вещество. При температуре выше 200є C разлагается с образованием так называемых карамелей. Сахароза не растворима в неполярных органических растворителях, в абсолютном метаноле и этаноле, умеренно растворима в атилацетате, анилине, в водных растворах метанола и этанола. Хорошо растворима в воде. Сахароза не обладает редуцентными свойствами, поэтому она устойчива к действию щелочей, но гидрализуется под влиянием кислот и ферментов сахараз с образованием D- глюкозы и D- фруктозы. Со щелочным металлами образует сахараты. Сахароза является одним из основных дисахаридов. Она гидролизуется HCl желудочного сока и сахаразой слизистой оболочкой тонкой кишки человека. Сахароза входит в состав сахара (99,75%), используемого для придания пище сладкого вкуса. Сахарозу также называют свекловичным сахаром. Другой представитель дисахаридов - лактоза (молочный сахар). Она состоит из остатков гелактозы и глюкозы. Лактоза - важная составная часть молока млекопитающих и человека. Образуется в процессе лактации в молочной железе из глюкозы и является для новорожденных ее источником. Лактоза облегчает всасывание кальция их кишечника. Содержание лактозы в женском молоке - 7 г/ 100 мл. В молоке коров и коз - 4,5г/100 мл. Мальтоза (солодовый сахар) — промежуточный продукт расщепления крахмала и гликогена в желудочно-кишечном тракте, происходящего под влиянием амилазы — фермента, выделяемого поджелудочной железой. Образующаяся мальтоза расщепляется затем мальтазой кишечного сока до двух остатков глюкозы. В свободном виде в пищевых продуктах мальтоза встречается в меде, солоде, пиве, патоке (мальтозной) и продуктах, изготовляемых с добавлением патоки (хлебобулочные, кондитерские изделия).
Целлобиоза — 4-(β-глюкозидо)-глюкоза, дисахаридов, состоящий из двух остатков глюкозы, соединённых β-глюкозидной связью; основная структурная единица целлюлозы. Белое кристаллическое вещество, хорошо растворимое в воде и в 45 — 48°-ном спирте. Плохо растворима в 96°-ном спирте и эфирах. Молекулярная масса = 342,30 г/моль. Температура плавления = 225 °C. Оптически активна, мутаротирует в растворе.
Для целлобиозы характерны реакции с участием альдегидной (полуацетальной) группы и гидроксильных групп, может образовывать гликозиды со спиртами, аминами, другими моносахаридами. Получают целлобиозу ферментативным гидролизом целлюлозы. В свободном виде целлобиоза содержится в соке некоторых деревьев. Целлобиоза образуется при ферментативном гидролизе целлюлозы бактериями, обитающими в желудочно-кишечном тракте жвачных животных. Затем целлобиоза расщепляется бактериальным ферментом β-глюкозидазой (целлобиазой) до глюкозы, что обеспечивает усвоение жвачными целлюлозной части биомассы. Рафиноза (раффиноза) — невосстанавливающий трисахарид, состоящий из остатков D-галактозы, D-глюкозы и D-фруктозы. Бесцветное растворимое в воде вещество с температурой плавления 80 °C (пентагидрат) и 119—120 °C (безводная). Один из распространённых растительных резервных углеводов (сахарная свёкла, семена хлопчатника и др.). Полное название: Альфа-D-Галактопиранозил-(1-6)-альфа-D-глюкопиранозил-(1-5)-Бета-D-фруктофуранозид. Мальтотриоза C18H32O16 восстанавливающий трисахарид, углевод, состоящий из трёх остатков моносахаридов, связанных гликозидными связями. Молекулярная масса 504.44 Паноза хорошо растворима в воде, образует тонкие призмоподобные кристаллы.Панозу можно легко выделить из реакционной смеси после удаления ферментируемых Сахаров и очистить осаждением основным ацетатом свинца и обработкой ионообменной смолой. Этот трисахарид легко кристаллизуется в виде а-с-аномера, кристаллы которого диморфны.
37.Специфичность действия ферментов, гидролизующих олигосахариды. Инверсия сахарозы. 37) Специфичность действия ферментов, гидролизирующих олигосахариды Одно из наиболее поразительных свойств ферментов их специфичность. Специфичность ферментов проявляется по-разному и может быть выражена в разной степени. Прежде всего, следует различать специфичность по отношению к субстрату и к типу химической реакции, катализируемой ферментом.Специфичность по отношению к типу химической реакции.Каждый фермент катализирует одну химическую реакцию или группу реакций одного типа. Наиболее ярким проявлением этого вида специфичности могут служить довольно частые случаи, когда одно и то же химическое соединение выступает как субстрат действия нескольких ферментов, причём каждый из них катализирует специфическую для него реакцию, приводит к образованию совершенно различных продуктов.У ферментов различают абсолютную, относительную и стереохимическую специфичность.Абсолютная специфичность – избирательная способность фермента катализировать только единственное из возможных превращений одного субстрата. Это можно объяснить конформационной и электростатической комплементарностью молекул субстрата и фермента. Например, фермент аргиназа катализирует только гидролиз аминокислоты аргинина, фермент уреаза – только расщепление мочевины и не действуют на другие субстраты.Относительная специфичность – избирательная способность фермента катализировать однотипные превращения сходных по строению субстратов. Такие ферменты оказывают воздействие на одинаковые функциональные группы или на один и тот же тип связей в молекулах субстратов. Так, например, разные гидролитические ферменты действуют на определённый тип связей: амилаза – на гликозидные связи; пепсин и трипсин – на пептидные связи; липаза и фосфолипаза – на сложноэфирные связи. Действие этих ферментов распространяется на большое число субстратов, что позволяет организму обойтись малым количеством пищеварительных ферментов - иначе их потребовалось бы намного больше.Стереохимическая (оптическая) специфичность - избирательная способность фермента катализировать превращение только одного из возможных пространственных изомеров субстрата. Так, большинство ферментов млекопитающих катализирует превращение толькл L-изомеров аминокислот, но не D-изомеров. ферменты, участвующие в обмене моносахаридов, наоборот, катализируют превращение только D-, но не L-фосфосахаров. Гликозидазы специфичны не только к моносахаридному фрагменту, но и характеру гликозидной связи. Например, α-амилаза расщепляет α–1,4-гликозидные связи в молекуле крахмала, но не действует на α–1,2-гликозидные связи в молекуле сахарозы. Инверсия сахарозы Инверсия — гидролитическое расщепление сахарозы на глюкозу и фруктозу. Под влиянием кислот сахароза в водном растворе, присоединяя воду, распадается на фруктозу и глюкозу. Ион водорода кислоты действует при этом как катализатор. Полученная смесь глюкозы и фруктозы вращает плоскость поляризации уже не вправо, как сахароза, а влево, так как левая вращательная способность образовавшейся фруктозы больше правого вращения глюкозы; происходит превращение правого вращения в левое, вследствие чего и реакция эта названа инверсией сахарозы. Реакция инверсии: С 12 Н 22 О 11 + Н 2 О = С 6 Н 12 О 6 + С 6 Н 12 О 6. Реакция — бимолекулярная, т. е. идет между двумя молекулами: сахар и вода. Но если рассматривать реакцию инверсии в разбавленных водных растворах, то количество воды будет мало изменяться, так как лишь незначительная часть ее будет связываться с сахарозой. Меняться постепенно будет только количество сахарозы, а реакция, таким образом, будет идти как мономолекулярная и необратимая (глюкоза с фруктозой обратно под влиянием кислоты не соединяется). Законы таких реакций были изучены именно путем наблюдения инверсии (Вильгельми). Так как при нормальной работе сахарного завода никогда не допускают значительной инверсии сахарозы, то концентрацию сахарозы можно считать почти постоянной: лишь ничтожно малая доля этой концентрации затрачивается на образование инвертного сахара. Поэтому можно здесь при вычислении накопившегося инвертного сахара обойтись без обычного логарифмического уравнения мономолекулярной реакции. По закону действующих масс количество образовавшегося инвертного сахара должно быть пропорционально начальной концентрации сахарозы (действующая масса) и времени
38) Лактоза и ее превращение в организме человека. Лактоза (от лат. lactis — молоко) C12H22O11 — углевод группы дисахаридов, содержится в молоке и молочных продуктах. Молекула лактозы состоит из остатков молекул глюкозы и галактозы.Лактозу иногда называют молочным сахаром. Такое название ей дал шведский химик – первооткрыватель огромного количества органических и неорганических веществ Карл Вильгельм Шееле в 1780 году. Он же внес ее в ряд углеводов под названием «лактоза». Впервые же выделил лактозу приблизительно на 160 лет раньше итальянский исследователь Фабрицио Бартолетти.Типичная черта лактозы — плохая усвояемость в организме взрослого человека. Если в организме детей лактоза в желудочно-кишечном тракте под влиянием фермента «лактазы» расщепляется и всасывается почти моментально, то во взрослом организме она в нерасщепленном виде проходит до самого толстого кишечника. В кишечнике лактоза начинает бродить с образованием большого количества токсинов и газов.
Плохое расщепление лактозы связано с тем, что в организме взрослого человека такой фермент, как «лактаза», практически отсутствует. И усвояемость молока зависит от того, в какой мере другие ферменты пищеварительной системы возьмут на себя роль «лактазы». У тех, у кого этого не происходит совсем, устанавливается стойкая непереносимость к молоку и молочным продуктам.В кисломолочных продуктах лактоза уже разрушена бактериями молочного брожения, поэтому они и усваиваются так легко даже в организме взрослого человека. Лактоза - дисахарид, образованный остатками D-галактозы и D-глюкозы, по современной номенклатуре углеводов относится к классу олигосахаридов, а именно дисахаридов (биоз).Лактоза содержит 12 связанных атомов углеродов 22 атома водорода, 9 гидроксильных атомов, 1 эфирный и 1 карбоксильный. Лактоза может синтезироваться химическим и биологическим путем.Теоретический химический синтез лактозы может быть осуществлен по равенствуС6Н12О6 + С6Н12О6 С12Н22О11 + Н2Оглюкоза галактоза лактоза водаМеханизм биологического образования лактозы в организме лактирующего животного до конца еще не выяснен. Если предложить, что лактоза синтезируется в организме, то единственным источником синтеза ее является глюкоза крови, приносимая к вымени. Глюкоза пространственной перестройкой (галактозогенезом) превращается в галактозу. В молоке, кроме лактозы, содержится другие углеводы и их производные. Из моносахаридов (моноз) молока важное значения имеет глюкоза и галактоза, являющиеся структурными элементами молекулы лактоза и ее гидролиза.
39.Гликозилтрансферазы и биосинтез полисахаридов. Роль НДФС в биосинтезе полисахаридов.Все разнообразие структур природных полисахаридов - результат трех типов биосинтетических процессов. Первым из них служит последовательный перенос отдельных моносахаридных остатков от нуклеотидсахаров на растущую цепь с участием специфических ферментов гликозилтрансфераз, обеспечивающих необходимое положение истереохимию образующейся гликозидной связи; таким способом синтезируются как монотонные последовательности моносахаридных остатков в го могликанах, так и лишенные признаков регулярности гетерополисахаридные цепи гликопротеинов.Второй тип - сборка олигосахаридного "повторяющегося звена" по первому типу реакций и его последующая полимеризация с образованием строго регулярных полимерных молекул, характерных для полисахаридных цепей липополисахаридов грамотрицательных бактерий или для бактериальных капсульных полисахаридов.Наконец, полисахариды, построенные по первому или второму типу, могут испытывать постполимеризац. модификации (третий тип биосинтеза), которые включают замещение атомов Н гидроксильных групп на ацильные остатки (ацетилирование, сульфатирование), присоединение боковых моно- и олигосахаридных остатков и даже изменение конфигурации отдельных моносахаридных звеньев (таким путем в результате эпимеризации при атоме С-5 образуются остатки L-гулуроновой кислоты из D-маннуроновой в составе альгинатов, а также остатки L-идуроновой кислоты из D-глюкуроновой в составе мукополисахаридов). Последние реакции часто приводят к нарушению (маскировке) первоначальной регулярности цепей полисахаридов и к образованию нерегулярных или блочных структур. Роль НДФС в биосинтезе полисахаридов нуклеозиддифосфатсахара (НДФС). С их помощью происходит биосинтез полисахаридов, таких как крахмал и гликоген, широко распространенных в природе дисахаридов сахарозы и лактозы и многих других углеводов, а также внутримолекулярные превращения сахаров – эпимеризация, окисление и восстановление. Так, под действием эпимеразы из УДФ-глюкозы образуется УДФ-галактоза, а под действием дегидрогеназы – УДФ-глюкуроновая кислота. Биологическая роль НДФС была выяснена в конце1950-х годов Луисом Лелуаром, который получил за это открытие Нобелевскую премию. Идея заключалась в том, что НДФС могут участвовать в ферментативных реакциях только в опредеделённых конформациях, и если в результате модификации аналог НДФС не может принимать такую конформацию, то он будет неспособен вступать в реакцию.
40.Полисахариды второго порядка. Общая характеристика. Крахмал и гликоген как запасная форма полисахаридов. Структурная организация. 40. Полисахариды - это высокомолекулярные углеводы, состоящие из большого числа моносахаридов. Их молекулярная масса велика, от нескольких тысяч до нескольких миллионов дальтон. Как и предыдущая группа углеводов, полисахариды второго порядка могут гидролизоваться до моносахаридов. Крахмал имеет структуру, представленную двумя формами полисахаридов: амилозой и амилопектином. В функциональном отношении различают полисахариды резервного и структурного назначения. Типичные резервные полисахариды - крахмал и гликоген. К структурным полисахаридам относят клетчатку (целлюлозу). Крахма́л (C6H10O5)n — полисахариды амилозы и амилопектина, мономером которых является альфа-глюкоза. Крахмал, синтезируемый разными растениями в хлоропластах, под действием света при фотосинтезе, несколько различается по структуре зёрен, степени полимеризации молекул, строению полимерных цепей и физико-химическим свойствам.Гликоген — (C6H10O5)n, полисахарид, образованный остатками глюкозы; основной запасной углевод человека и животных. Гликоген является основной формой хранения глюкозы в животных клетках. Откладывается в виде гранул в цитоплазме во многих типах клеток (главным образом печени и мышц). Гликоген образует энергетический резерв, который может быть быстро мобилизован при необходимости восполнить внезапный недостаток глюкозы. Гликогеновый запас, однако, не столь ёмок в калориях на грамм, как запас триглицеридов (жиров). В качестве запасного углевода гликоген присутствует также в клетках грибов.
41.Амилазы - распространение в природе и механизм действия. Роль амилаз в пищевой промышленности.
41. Амилаза — фермент, гликозил-гидролаза, расщепляющий крахмал до олигосахаридов, относится к ферментам пищеварения. Именно амилаза приводит к появлению сладковатого вкуса при длительном пережёвывании крахмалосодержащих продуктов (например, из риса или картофеля), но без добавления сахара. Амилаза присутствует в слюне (птиалин), где начинает процесс пищеварения. Амилазы очень широко распространены в природе. Они синтезируются многими микроорганизмами (бактерии, грибы, актиномицеты, дрожжи), животными и растениями. До развития ферментной промышленности главным промышленным источником получения амилаз в европейских странах было проросшее зерно (солод). Для медицинских целей амилазы получали из животного сырья. В настоящее время главным источником амилаз являются микроорганизмы, особенно бактерии, грибы и реже дрожжи. В современных технологиях амилаза используется как одно из важных составляющих специальной добавки, ускоряющий процесс брожения. Бактериальная амилаза используется в стиральных порошках для разложения крахмала, присутствующего в белье.Механизм действия. К группе амилотических ферментов относятся a- и b-амилазы, глюкоамилаза, пуллуланаза, изоамилаза и некоторые другие ферменты. Амилазы бывают двух типов: эндо- и экзоамилазы. Четко выраженной эндоамилазой является а-амилаза, способная к разрыву внутримолекулярных связей в высокополимерных цепях субстрата.При изучении механизма действия амилаз имеются определенные сложности, и прежде всего они заключаются в том, что субстрат - крахмал неоднороден и имеет различные характеристики по степени полимеризации гли-козидной цепи и количеству ветвлений.В пищевой промышленности зарегистрирована в качестве пищевой добавки E1100 как улучшитель муки и хлеба.
42.Фосфорилазы. Взаимопревращение крахмала и сахарозы в растениях. 42. Фосфорилазы - ферменты класса трансфераз. Катализируют обратимые реакции переноса гликозильных групп (остатков моносахаридов) на ортофосфат (фосфоролиз). Фосфорилазная реакция может быть представлена уравнением: А – Г + Ф А + Г – Ф, где Г – гликозильная группа, А – акцептор гликозильной группы, Ф – ортофосфат. Фосфорилазы имеют универсальное распространение в природе, встречаются у простейших, в животных и растительных тканях. Играют важную роль в живых организмах, катализируя ключевые реакции метаболизма, связанные с использованием запасных углеводов, а следовательно, с обеспечением клеток энергией. В растении происходит процесс взаимопревращение крахмала в сахар и обратно.
Превращение крахмала в сахар — патоку — можно проводить двумя путями, и оба легко могут быть осуществлены домашними средствами. Один путь — это превращение под действием фермента на крахмал. Такой фермент легко получить из прорастающих семян. Другой путь — нагревание с кислотой — применяется в фабричном производстве патоки из крахмала.
43.Клетчатка и гемицеллюлозы, их свойства и ферментативный гидролиз.
| Гемицеллюлозы (полу клетчатки) — полисахариды, которые вместе с целлюлозой входят в состав оболочек растительных клеток и отличаются от нее тем, что легче гидролизуются. В их состав входят пентозаны, образующие при гидролизе пентозы (арабинозу ксилозу), гексозаны, гидролг зующиеся до гексоз (манноз, галактоза, глюкоза, фруктоза и группа смешанных полисахаридов, гидролизующихся до пентоз, гексоз и уроновых кислот. Гемицеллюлозы обычно имеют разветвленное строение; порядок расположения моноз внутри полимерной цепи неодинаков. Связь их Друг с другом осуществляется с участием полуацетального гидроксила и гидроксильных групп у 2, 3, 4, 6-го углеродных атомов. Они растворяются в щелочных растворах. Кислотный гидролиз гемицеллюлозы протекает значительно легче, чем целлюлозы. К гемицеллюлозам относятся гексозаны — галактаны и маннаны, которые при гидролизе дают галактозу и маннозу, и пектозаны — арабан и ксилан, при гидролизе которых образуются арабиноза и ксилоза. Для растений гемицеллюлозы выполняют не только роль строительного материала (пентозаны), но и запаса питательных веществ (гексозаны). Они находятся в плодах и ягодах (0,55—4,25%), в зерне пшеницы и ржи (7—8%), ячмене и просе (10—11%) и др. Целлюлоза (клетчатка) широко распространена в растениях, составляя основу клеточных стенок. Молекула целлюлозы состоит из остатков глюкозы, число которых колеблется от 3000 до 10 000, составленных в виде неразветвленной цепи. Целлюлоза обладает большой механической прочностью, что обусловливается ее строением. Молекулы целлюлозы вытянутой нитевидной формы соединены в пучки (мицеллы) водородными мостиками, а мицеллы объединены в волокна.Целлюлоза, которая находится в мякоти плодов и овощей, мучнистом ядре зерна злаков (до 0,6%), в пище человека желательна, так как она усиливает перистальтику кишечника и способствует передвижению пищи. В оболочках зерна и семян, кожице плодов и овощей находится одревесневшая целлюлоза, в которой пустоты между мицеллами заполнены инкрустирующими веществами (лигнином, кутином), что придает ей особую прочность. Целлюлоза содержится в пищевых продуктах в небольших количествах: в плодах и овощах — 0,5—1,6%, пшене — 0,8, зерне пшеницы — 2,0, зерне ячменя — 4,5% и т. д. Целлюлоза нерастворима в воде, органических растворителях, слабых кислотах и щелочах. Это свойство целлюлозы используется при количественном определении ее в пищевых продуктах. Она растворима в реактиве Швейцера — аммиачном растворе гидрата окиси меди. При гидролизе целлюлозы концентрированными кислотами и ферментами целлюлозами (широко распространенными у микроорганизмов) образуется глюкоза. Для организма человека целлюлоза питательной ценности почти не имеет, так как нет ферментов для ее расщепления до Сахаров. Только незначительная часть ее подвергается расщеплению ферментами микроорганизмов, находящимися в кишечнике. При повышенных температурах при гидролизе образуется в качестве конечного продукта D-глюкоза. В ходе гидролиза постепенно идет деполимеризация крахмала и образование декстринов, затем мальтозы, а при полном гидролизе глюкозы. Деструкция крахмала, которая начинается с набухания и разрушения крахмальных зерен и сопровождается его деполимеризацией (частичной или более глубокой) до образования в качестве конечного продукта глюкозы, происходит при получении многих пищевых продуктов - патоки, глюкозы, хлебобулочных изделий, спирта и т. д. |
44.Пектиновые вещества, их свойства, ферментативные, роль в пищевых технологиях и в питании человека. Влияние типа связей в полисахаридах на степень их усвоения в организме животных и человека.44. Пектиновые вещества — это высокомолекулярные углеводы растительного происхождения, главным структурным компонентом которых является D-галактуроновая кислота; встречаются в тканях наземных растений и в некоторых водорослях. В тканях некоторых растений содержание пектиновых веществ достигает 30% сухого веса (например, в белой части кожуры цитрусов), в других — их содержание не превышает долей процента. К пектиновым веществам относятся:
-пектовая кислота, которая построена из остатков D-галактуроновой кислоты, связанных α-1,4-гликозидными связями в длинные цепи (является основой всех пектиновых веществ);- пектаты — соли пектовой кислоты;- пектины — продукты различной степени метилирования пектовой кислоты по карбоксильным группам, растворяются в воде с образованием плотных гелей;- пектинаты — соли пектинов;- протопектины — нерастворимые в воде вещества высокого молярного веса, в которых линейные молекулы пектинов связаны поперечными мостиками, и другие производные пектина.
Главные характерные свойства пектиновых соединений. Пектиновые вещества, являясь одним из компонентов растительной клетки, регулируют ее тургор. Они обеспечивают растениям характерную для них влажность. Пектины относят к растворимым пищевым волокнам. Они в основном содержатся в покровной ткани плодов. Основным сырьем в Венгрии, в котором находится пектин – являются яблоки. Пектиновые вещества локализуются в их кожуре. Основные свойства пектинов В природе пектин содержится в нерастворимой форме – в виде протопектина. Извлекают пектин в промышленных условиях из выжимок яблок, кожуры крупноплодных цитрусовых (апельсин, грейпфрутов, мандарин, лимонов), жома сахарной свеклы. Производят его в виде порошка и концентрата. Пектиновый порошок не имеет запаха, цвет изменяется от светло-кремового до светло-бежевого в зависимости от вида сырья. В воде при растворении образует коллоидный раствор. Пектиновый концентрат представляет собой вязкую непрозрачную жидкость с запахом свойственным исходному сырью.Для пектина характерны два основных свойства - комплексообразующая и студнеобразующая способности, используемые в зависимости от области их применения.
Роль пектинов в питании Пектин входит в состав всех зеленых растений, поэтому является неотъемлемой частью нашей пищи. Последние исследования выявили, что:а) когда пектин попадает в желудочно-кишечный тракт, он образует гели, которые, по мере продвижения по кишечнику, захватывают токсичные вещества и, таким образом, защищают слизистые от раздражения;б) в процессе усвоения пищи пектин соединяется с токсинами, солями тяжелых металлов и радионуклидами, образуя нерастворимые комплексы, которые не могут всасываться в слизистую оболочку желудочно-кишечного тракта, поэтому легко выводятся из организма;в) пектины улучшают работу (перистальтику) кишечника, способствуя более быстрому выведению токсинов и недоокисленных веществ из организма человека;г) попадая в кишечник, пектиновые вещества оказывают бактерицидное действие на болезнетворные микробы, поскольку сдвигают рН его среды в более кислую сторону;д) связывая желчные кислоты, пектины предупреждают атеросклероз
Пектин происходит в той или иной степени во всех растениях и плодах.Он не находится в животных тканях.Но его много в фруктах и овощах.
45.Классификация липидов. Жиры, их свойства, ферментативный гидролиз.45. Различают три основных группы липидов: нейтральные липиды, фосфолипиды и гликолипиды. В организмах встречаются также отдельные группы минорных липидов (фосфатидилглицерин, липопептиды, диольные липиды и др.). Нейтральные липиды. Среди нейтральных липидов наиболее распространены жирные кислоты. В настоящее время известно свыше 800 природных жирных кислот, которые условно можно разделить на три группы: насыщенные (пальмитиновая, стеариновая и др.),моноеновые (олеиновая, эруковая и др.) и полиеновые (линолевая, линоленовая, арахидоновая, эйкозапентаеновая, докозагексаеновая и др.) Наиболее богато пальмитиновой кислотой (более 50%) пальмовое масло. Она встречается также в животных жирах и хлопковом масле (25%). Стеариновой кислоты обычно в жирах не более 10%. Исключением является бараний жир, в котором ее более 30%. Помимо этих двух насыщенных жирных кислот в природе достаточно широко распространены лауриновая и миристиновая кислоты. Олеиновой кислоты больше всего в оливковом и салатном подсолнечном масле (около 80%). В других жирах и маслах ее содержится от 5 до 40%. В маслах из семян горчицы и рапса до 50% другой моноеновой жирной кислоты - эруковой. Главной составляющей многих растительных масел (подсолнечного, соевого, кукурузного, хлопкового) является линолевая кислота, ее содержание в них составляет 50-70%. В льняном масле больше всего линоленовой кислоты. Жиры рыб и других морских животных богаты полиеновыми: эйкозапентаеновой и докозагексаеновой. Арахидоновая кислота входит в состав фосфолипидов млекопитающих. Все полиеновые жирные кислоты являются обязательными компонентами фосфолипидов биомембран.В группу нейтральных липидов входят также триглицериды, воски, эфиры стеринов, N-ацилэтаноламиды жирных кислот и церамиды, состоящие из сфингозина (2-амино-4-октадекен-1,3-диола) и жирной кислоты. Воски и эфиры стеринов имеют общую химическую формулу. Наиболее известны воски - продукты деятельности насекомых (пчелиный), животных (ланолин) и растений. Триглицериды являются основным веществом всех жиров и масел. N-ацилэтаноламиды и церамиды – ценные биологически активные вещества. В первую очередь это относится к этаноламидам жирных кислот. Фосфолипиды – это сложные липиды, содержащие фосфорную кислоту. Они содержатся во всех живых клетках, являются важнейшими компонентами биологических мембран нервной ткани. В составе липопротеидов крови участвуют в транспорте жиров, жирных кислот и холестерина. Фосфолипиды довольно широко используются в пищевой и фармацевтической промышленности.Фосфатидилхолин - главный фосфолипид большинства типов животных. Его содержание обычно составляет не менее 50% суммы фосфолипидов. Вторым по значению фосфолипидом у животных обычно является фосфатидилэтаноламин. В большинстве бактерий фосфатидилхолина нет, а более 60-70% их фосфолипидов составляет фосфатидилэтаноламин. Оба липида присутствуют в большинстве растений, для этих организмов очень важен фосфатидилглицерин. Это единственный фосфолипид синезеленых водорослей, главный фосфолипид фотосинтетического аппарата всех растений. Сфингомиелин является важным компонентом клеток эволюционно продвинутых типов животных. В эритроцитах некоторых млекопитающих, в частности овец, он заменяет фосфатидилхолин в качестве главного фосфолипида. Заслуживают упоминания и несколько других фосфолипидов: фосфатидилинозит, дифосфатидилглицерин (кардиолипин), фосфатидилсерин, фосфатидная кислота. Гликолипиды —— сложные липиды, образующиеся в результате соединения липидов с углеводами. Гликолипиды входят в состав клеточных мембран. Они широко представлены в тканях, особенно в нервной ткани, в частности в ткани мозга. Гликолипиды локализованы преимущественно на наружной поверхности плазматической мембраны, где их углеводные компоненты входят в число других углеводов клеточной поверхности. Гликолипиды растений играют важную роль в процессе фотосинтеза. Главной формой гликолипидов в животных тканях являются гликосфинголипиды. Они содержат церамид, состоящий из сфингозина (2-амино-4-октадекен-1,3-диола) и жирной кислоты, а также один или несколько остатков сахаров. Двумя простейшими соединениями этой группы являются галактозилцерамид и глюкозилцерамид. Галактозилцерамид — главный гликосфинголипид мозга и других нервных тканей, но в небольших количествах он встречается и во многих других тканях. Простые гликосфинголипиды в тканях, отличных от нервной, представлены главным образом глюкозилцерамидом; в небольших количествах он имеется и в ткани мозга.Гликосфинголипиды, являющиеся компонентами наружного слоя плазматической мембраны, могут участвовать в межклеточных взаимодействиях и контактах. Некоторые из них являются антигенами, например антиген Форссмана и вещества, определяющие группы крови. Сходные олигосахаридные цепи обнаружены и у других гликопротеинов плазматической мембраны.
| Жиры — органические соединения, входящие в состав животных и растительных тканей и состоящие в основном из триглицеридов (сложных эфиров глицерина и различных жирных кислот). Помимо триглицеридов, в состав жиров входят вещества, обладающие высокой биологической активностью: фосфатиды, стерины, витамины. Смесь различных триглицеридов составляет так называемый нейтральный жир. Жиры и жироподобные вещества объединяют обычно под названием липиды.Жиры – одна из основных групп веществ, входящих, наряду с белками и углеводами, в состав всех растительных и животных клеток. В организме животных различают запасные и плазматические жиры. Запасные жиры откладываются в подкожной клетчатке и в сальниках и являются источником энергии.Плазматические жиры структурно связаны с белками и углеводами и входят в состав большинства мембран. Жиры обладают высокой энергетической ценностью. Благодаря низкой теплопроводности жиры играют важную роль в теплорегуляции животных организмов, предохраняя животных, особенно морских, от переохлаждения. Вследствие своей эластичности жирыиграют защитную роль в коже позвоночных и в наружном скелете насекомых. Жиры – необходимая составная часть пищи.Гидролиз жиров |
| Гидролиз жиров протекает при наличии в них воды. При обычной температуре скорость гидролиза ничтожна, так как в жире растворено мало воды (десятые доли процента). Ускоряется гидролиз жира под действием катализаторов, как-то: фермента липазы, щелочей, кислот. Усиливается гидролиз жира при повышении температуры, давления и др. Ферментативный гидролиз жира возникает под действием ферментов плесеней на поверхности соприкосновения жира и воды. Степень расщепления жира в этом случае пропорциональна поверхности контакта. |
46.Липазы. Бета – окисление жирных кислот. Коэнзим А и его роль в процессе обмена. Липоксигеназа и ее роль в пищевой промышленности.. ЛИПАЗЫ (от греч. lipos - жир) (триацилглицерол - ацилгидролазы), ферменты класса гидролаз, катализирующие гидролиз эфиров глицерина и высших карбоновых к-т.Иногда к липазам относят также фермент холестеролэстеразу, расщепляющую эфиры холестерина. Гидролиз триацилглицеринов идет преимущественно в положении 1. Ди- и моноацилглицерины гидролизуются медленно. Липазы широко распространены у млекопитающих (панкреатич. и тканевые липазы), растений, дрожжей и бактерий. Из плазмы крови и молока выделена также липаза (липопротеидлипаза), гидролизующая триацилглицерины, связанные с белком, которые входят в состав липопротеинов низкой плотности. Наиболее изучена панкреатичная липаза свиньи. Она существует в виде 2 форм (А и В) и представляет собой гликопротеин. Состоит из одной полипептидной цепи с 6 дисульфидными связями и содержит 2 меркаптогруппы, блокирование которых не влияет на активность фермента. Активный центр этого фермента содержит серин (он занимает положение 110 в полипептидной цепи). В организме панкреатич. липаза функционирует в комплексе с колипазой (белок с мол. м. 10 тыс.), по-видимому защищающей липазу от ингибирующего действия солей желчных к-т. Холестеролэстераза обладает гидролитич. и синтетазной активностью. Этот фермент (мол. м. 65-69 тыс.) из панкреатич. сока крысы проявляет оптимальную активность при рН 6-7. Особенность липаз - их активация на повседневности раздела фаз, образованных липидом и водой. Фермент очень медленно гидролизует эфиры глицерина и карбоновых кислот с короткой углеводородной цепью, находящиеся в истинных растворах. Однако при увеличении концентрации эфира выше критич. концентрации мицеллообразования скорость гидролиза резко возрастает. Предполагают, что липазы имеют специфич. участок, ответственный за "активацию пов-стью", а сама активация обусловлена конформац. изменениями молекулы фермента. Оптим. каталитич. активность липаз проявляется при рН 8-9. Однако нек-рые липазы растений и микроорганизмов активны в слабокислой среде (рН 4-6). Липазы играют важную роль в обмене липидов. Их используют для обезжиривания шкур, ароматизации и ускорения созревания сыров, как компонент лек. ср-в. β-окисление жирных кислот После попадания в клетки жирные кислоты активируются путем образования ацил-КоА Для этого нужны две богатые энергией ангидридные связи АТФ. В матрикс митохондрий активированные жирные кислоты попадают в виде ацилкарнитина, который является трансмембранным переносчиком.Деградация жирных кислот происходит в митохондриальном матриксе путем окислительного цикла реакций, при котором последовательно отщепляются С2-звенья в виде ацетил-КоА (активированной уксусной кислоты). Последовательное отщепление ацетильных групп начинается с карбоксильного конца активированных жирных кислот каждый раз между С-2 (α-атомом) и С-3 (β-атомом). Поэтому цикл реакций деградации называется β-окислением. Пространственно и функционально β-окисление тесно связано с цитратным циклом и дыхательной цепью Первая стадия β-окисления — дегидрирование активированной жирной кислоты (ацил-КоА) с образованием β-ненасыщенной жирной кислоты с двойной связью в транс -конфигурации (реакция дегидрирование). При этом оба атома водорода с электронами переносятся от фермента на электронпереносящий флавопротеин (ETF). ETF-дегидрогеназа переносит восстановительные эквиваленты на убихинон (кофермент Q), который является составной частью дыхательной цепи. Вторая стадия деградации жирной кислоты состоит в присоединении молекулы воды к двойной связи ненасыщенной жирной кислоты (реакция: гидратирование). На третьей стадии происходит окисление гидроксильной группы при С-3 в карбонильную группу (реакция: дегидрирование). Акцептором для восстановительных эквивалентов является НАД+ который передает их в дыхательную цепь. На четвертой стадии активированная β-кетокислота расщепляется ацилтрансферазой (β-кетотиолазой) в присутствии кофермента А: тиолитическое расщепление). Продуктами реакции являются ацетил-КоА и активированная жирная кислота, углеродная цепь которой короче на два углеродных атома по сравнению с длиной цепи исходной жирной кислоты.Для полной деградации длинноцепочечной жирной кислоты цикл должен многократно повторяться; например, для стеарил-КоА (18:0) необходимы восемь циклов. Образующийся ацетил-КоА может переноситься на оксалоацетат с образованием цитрата, промежуточного метаболита цитратного цикла. При избытке ацетил-КоА в печени образуются кетоновые тела. Коэнзим АЭнергия в чистом виде Об этом веществе современные физиологи и биохимики говорят с восторгом. Чем больше исследователи узнают об этом чуде природы, тем больше новых открытий оно сулит людям. Коэнзим А принимает самое активное участие в производстве энергии в каждой из 70 триллионов клеток нашего организма и необходим для любого вида мышечной деятельности. Даже когда вы утром открываете глаза, на это расходуются миллионы молекул коэнзима А. Если вы чувствуете себя вялым и уставшим, но хотите взбодриться, то вам надо повысить концентрацию коэнзима А в организме на 50%. И произойдет чудо! Вы почувствуете себя заново родившимся.Основным действующим началом и ядром молекулы коэнзима А является пантетин, получаемый из пантотеновой кислоты (витамин группы В). Его превращение в молекулу коэнзима А происходит молниеносно. Если сердце получает недостаточно кислорода (например, когда мы сидим без движения в кресле у телевизора), содержание пантетина в сердечной мышце падает до угрожающего минимума. А ведь пантетин (или коэнзим А) играет главную роль в снижении концентрации липидов, сжигая опасный жир. Коэнзим А снижает содержание холестерина и безостановочно гонит жир в топку мышечных клеток. Среди всех веществ, снижающих уровень липидов - это самое безопасное и наименее токсичное. Его токсичность практически равна нулю. Если вы чувствуете себя вялым и уставшим, но хотите взбодриться, то вам надо повысить концентрацию коэнзима
Липоксигеназа липоксидаза, фермент класса оксидоредуктаз (См. Оксидоредуктазы); окисляет ненасыщенные жирные кислоты до перекисей. Впервые найден в 1934 в соевых бобах; обнаружен также в семенах др. растений, мышцах рыб и млекопитающих. Основные субстраты Л. — линолевая, линоленовая, арахидоновая кислоты и их эфиры. Л. активируется полипептидами.
47.Прогорание жиров. Токсичность продуктов окисления жирных кислот. изменение состава жиров (См. Жиры) при хранении, в результате которого они становятся непригодными для пищи, приобретают крайне неприятный, «прогорклый» запах и вкус; это обусловливается появлением в их составе некоторых летучих альдегидов, кетонов и кислот невысокой молекулярной массы, а также нелетучих веществ перекисного характера. Различают два типа П. ж.: биохимическое, обусловленное действием плесеней, развивающихся на жире, и чисто химическое, связанное с окислением кислородом воздуха радикалов жирных кислот, входящих в состав глицеридов. Для предупреждения прогоркания производство жиров осуществляют в условиях, исключающих возможность заражения спорами плесени; жиры подвергаются тщательной очистке. Для предотвращения биохимического прогоркания вводят консервирующие вещества (например, соль). Хранить жиры (сливочное масло, маргарин и др.) следует в хорошо закупоренной таре при температуре от 2 до 6 °С.
48.Фосфатиды, их участие в построении биологических мембран и роль в пищевой промышленности.
48. ФОСФАТИДЫ, фосфолипиды, сложные липиды, в молекулах которых присутствует остаток фосфорной кислоты. Фосфатиды — сложные эфиры фосфорной кислоты и глицерина или аминоспирта сфингозина, которые посредством эфирной или амидной связи соединены с остатками насыщенных и ненасыщенных жирных кислот. К важнейшим фосфатидам относятся: фосфатидилэтаноламины, фосфатидилхолины, фосфатидилглицерины, фосфатидилсерины, дифосфатидилглицерины (кардиолипин), фосфатидилинозиты. Входят в состав клеточных и субклеточных структур животных, растений и микроорганизмов. Наличие полярных и неполярных группировок в молекулах фосфатидов обусловливает своеобразие физико-химических свойств и специфическую роль фосфатидов в построении и функционировании биологических мембран. Основную роль в биосинтезе фосфатидов выполняют цитидиновые нуклеотиды, с помощью которых строятся фосфодиэфирные связи. Биосинтез осуществляется преимущественно в печени, тканях кишечника, почках, мышцах, в меньших количествах — в мозге. Катаболизм фосфатидов протекает под действием липаз (фосфолипазы A1, A2, C, D), катализирующих специфическое расщепление сложноэфирных и фосфодиэфирных связей. Биохимическими исследованиями установлена роль фосфора и его различных производных в биосинтетических процессах, происходящих в растении. Обычно полагают, что в растениях фосфор содержится в виде следующих веществ: нуклеиновых кислот, эфиров фосфорной кислоты, не содержащих азота (фитин, гексозо - фосфорная кислота и глицерофосфорная кислота); неорганических фосфатов; фосфатидов.Нуклеиновые кислоты, неорганические фосфаты и эфиры глицерофосфорной, гексозофосфорной и инозитолфосфорной кислот сами по себе в масле не растворяются, но их присутствие в масле можно предполагать, так как они могут находиться в виде комплексов с другими веществами, растворимыми в масле. Фосфатиды локализуются в зрелых семенах преимущественно в гелевой части, а в масляной части могут находиться лишь в незначительных количествах. В основном они находятся в связанном состоянии с другими веществами семени (главным образом с белками и углеводами). Содержание фосфатидов в сырых маслах зависит от содержания их в перерабатываемых семенах, от соотношения фосфатиды: масло в данных семенах и от технологических условий - схемы и режима извлечения масла. Во всех случаях сырое масло отличается от нативного масла, содержащегося в клетках масличных семян. Это различие будет тем больше, чем интенсивнее воздействие технологических факторов в процессе переработки семян.
Фосфатиды являются полифункциональными соединениями, способными к различным взаимодействиям и превращениям; некоторые фосфатиды могут давать реакции, характерные для аминов и аминокислот, для жирных кислот и глицеридов. Фосфатиды обладают кислотными свойствами и могут взаимодействовать с веществами, имеющими щелочные функции, В определенных условиях фосфатиды могут взаимодействовать с металлами и белками.
49.Биосинтез жиров. 49 вопрос. Биосинтез жиров (жирных масел)
БИОСИНТЕЗ ЖИРОВ (ЖИРНЫХ МАСЕЛ). Состоит из неск. этапов и может быть представлен в виде общей схемы:
крахмал —> сахар —>глицерин —> жир {насыщенные кислоты } {ненасыщенные кислоты} Главные источники образования компонентов жира — гексозы, в первую очередь глюкоза и фруктоза. Иногда участвуют пентозы, маннит или др. продукты. Глицерин получается в процессе анаэробной диссимиляции углеводов путем восстановления глицеринового альдегида, образующегося из фруктозодифосфата под действием фермента альдолазы. Активный ацетат образуется окислением три-D-фосфоглицеральдегида. Биосинтез жирных кислот протекает в три фазы. Исходный материал — ацетил, активированный СоА. Процесс суммарно представлен на схеме (см. ниже). Малонил-СоА — важнейший промежуточный продукт при ферментативном синтезе жирных кислот. Это источник двууглеродного фрагмента, последовательно присоединяющегося при синтезе жирных кислот. Биотин является коферментом ацетил-СоА-карбоксилазы, который катализирует реакцию присоединения СО2- Включение ацетил-СоА в жирную кислоту происходит в митохондриях. Биосинтез триацилглицеринов (триглицеридов) локализован в микросомной фракции клеток. Один из основных этапов — образование L-глицерофосфата из свободного глицерина. Глицеролкиназа, катализирующая эту реакцию, обнаружена в растворимой фракции клетки.
50.Воски, стероиды и растворимые в жирах пигменты. Каротиноиды и эргостерол как провита

