 2015-07-04
2015-07-04 769
769Механізм передачі сигналу від рецептору всередину клітини
Передача сигналу від поверхні всередину клітини є необхідною для функціонування будь-якого організму: для дії гормонів, нейромедіаторів. просто для спілкування клітин одна з одною. Для цього природою розроблено спеціальні механізми. Таким чином, клітина спілкується з зовнішнім оточенням через велику кількість рецепторів на своїй поверхні. Всі позаклітинні сигнали отримуються зовнішньою поверхнею клітини і переносяться через мембрану з залученням трансмембранних рецепторних білків, які є інструментами в конвертації сигналу ззовні клітини у внутрішньоклітинні біохімічні події. Всередині клітини сигнал поширюється вздовж внутрішньоклітинних сигнальних шляхів, конвертується в різні біохімічні процеси (трансдукція сигналу) на різних рівнях організації клітини та підтримуються і значно ампліфікуються при досягненні пункту призначення. Принцип механізмів сигналінгу є універсальний, а деталі дозволяють закодувати і розшифрувати все різноманіття зовнішніх сигналів, що їх може отримати клітина.
Існує декілька загальних каскадів передачі сигналу. Вони складаються із спільних елементів:
І -рецептор, що сприймає сигнал на зовнішній поверхні клітини;
2- каскад реакцій, що призводить до фосфорилювання внутрішньоклітинних білків. Фосфорилювання-дефосфорилювання - універсальний механізм активації у еукаріотів (у прокаріотів таку функцію виконує метилювання);
3 - система вторинних месенджерів, що передає сигнал всередину клітини;
4 - активація певних генів, що відбувається внаслідок модифікацій і міграції у ядро активаційних ядерних факторів. В результаті цього клітина змінює свою генетичну програму, починає синтезувати нові білки, як то цитокіни, хемокіни, молекули клітинної адгезії, та інші поверхневі білки, проліферувати та диференціювати або навпаки включає механізм запрограмованої загибелі - тобто реагує на сигнал, що надійшов з-зовні.
1. Найбільш варіабільним елементом всієї системи є рецептор. Саме він розпізнає специфічний сигнал. Завдяки наявності великої кількості різноманітних рецепторів клітина здатна реагувати на все різноманіття зовнішніх стимулів. Клітина може регулювати свою готовність до сприйняття сигналу шляхом варіювання рівня експресії рецептору. Так, наприклад, при активації Т лімфоцитів, як уже зазначалось, вони одночасно починають синтезувати інтерлейкін-2 та рецептор до інтерлейкіну-2, тому що активована клітина повинна вміти розпізнати активаційний сигнал.
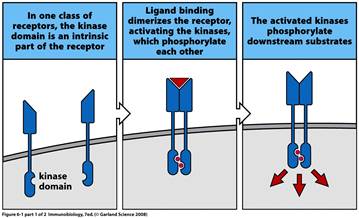
Рецептори - це, як правило, глікопротеїди, що мають позаклітинну, трансмембранну і цитоплазматичну частини. Всі рецептори можна розділити на дві великі групи: ті, що мають досить довгу цитоплазматичну ділянку, що здатна сама передатисигнал від рецептору (тобто має ферментативну активність), і ті, що мають коротку цитоплазматичну ділянку і для передачі сигналу потребують додаткових молекул, об'єднаних із самим рецептором в єдиний білковий комплекс. До першого типу рецепторів належать, наприклад, рецептори до інсуліну, фактору росту нервів, епідермального фактору росту,
 |
до другого - антиген-специфічні рецептори Т і В лімфоцитів.
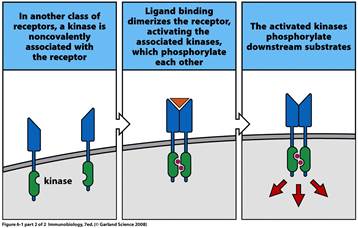 |
Важливим моментом для трансдукції сигналу через антиген-специфічні рецептори Т і В лімфоцитів є їх димеризація або кластеризація при взаємодії з антигеном, що приводить до активації ферментних комплексів та інших білків, асоційованих з рецептором.
2. Ферменти, які в основному асоційовані з рецепторною активацією, протеїнкінази. Це велика група ферментів, що каталізують ковалентне приєднання фосфатних груп до білків шляхом оборотного процесу, який отримав назву білкового фосфорилювання. Рецептор-асоційовані протеїнкінази в нормі неактивні, але після зв’язування ліганда з позаклітинною частиною рецептора вони активуються і передають сигнал далі, фосфорилюючи і активуючи інші сигнальні молекули всередині клітини.
У ссавців протеїнкінази фосфорилюють білки по трьом амінокислотним залишкам, а саме: тирозину, серину чи треоніну. В сигнальній трансдукції насьогодні вважають найбільш важливим тирозинове фосфорилювання.
Таким чином, процес фосфорилювання є ініаторним внутрішньоклітинним сигналом, активованим зв’язуванням ліганду з рецептором.
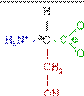
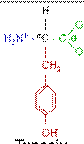 Тирозин (Tyr/Y) Серин (Ser/S) Треонін (Thr/T)
Тирозин (Tyr/Y) Серин (Ser/S) Треонін (Thr/T)
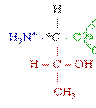 |
2. Системи месенджерів.
Необхідно зазначити, що вторинні месенджери мають ряд особливостей. По-перше, це-відносно невелика, порівняно з біополімерами Мм, оскільки треба з великою швидкістю дифундувати у цитоплазмі. По-друге, має дуже швидко розщеплюватись, а у випадку з кальцієм- відкачуватись, оскільки сигнальна система не може залишатись у ввімкненому стані тоді, коли дія сигналу вже закінчилась.








