 2015-07-04
2015-07-04 13433
13433За счет чего же возникает потенциал действия, какие механизмы лежат в его основе? Приоритет в ответах на эти вопросы принадлежит двум выдающимся ученым - А. Ходжкину и Э. Хаксли, проводившим исследования в 30-40-х годах прошлого века. Их достижения в детальном объяснении столь сложных биофизических молекулярных процессов трудно переоценить. Все последующие исследования по регистрации одиночных каналов возбудимых тканей лишь дополнили сформированные ими представления о молекулярных механизмах возбуждения.
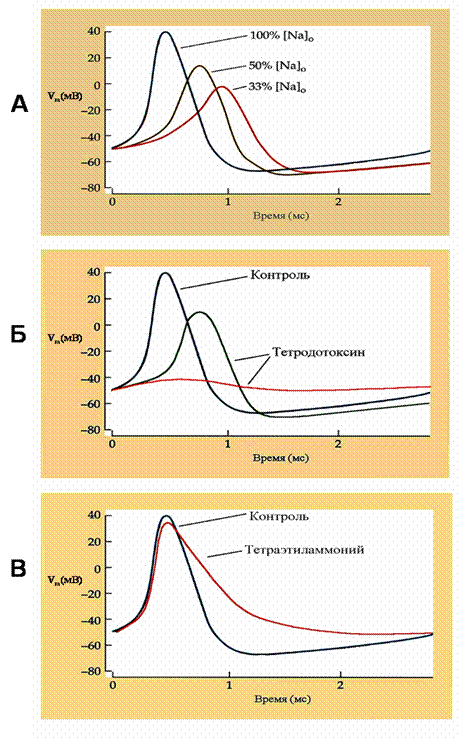
Рис. 19. Роль ионов натрия и калия в возникновении потенциала действия
Потенциалы действия зарегистрированы в гигантском аксоне кальмара. Изменения потенциала действия при уменьшении внеклеточной концентрации ионов натрия (А), при добавлении разных концентраций тетродотоксина – блокатора натриевых каналов (Б) и тетраэтиламмония - блокатора калиевых каналов (В).
Эти ученые работали на гигантском аксоне кальмара, очень удобном объекте, поскольку его диаметр более миллиметра. Крупные размеры позволяли менять внутриклеточный ионный состав аксоплазмы и вводить внутрь аксона один или несколько электродов для отведения потенциала действия и раздражения аксона. Были проведены исследования по влиянию изменений вне- и внутриклеточной концентрации ионов и природных токсинов, блокирующих различные ионные каналы, на форму потенциала действия (рис.19). Оказалось, что при снижении концентрации ионов натрия в окружающей среде потенциал действия уменьшается по амплитуде; следовательно, потенциал действия зависит от концентрации внеклеточного натрия (рис. 19А). При исследовании эффекта тетродотоксина, специфического блокатора натриевых каналов, был получен эффект, аналогичный снижению внеклеточной концентрации ионов натрия - потенциал действия уменьшался, а при высоких концентрациях токсина – не возникал (рис. 19Б). Было сделано заключение, что потенциал действия связан с открытием потенциал- активируемых натриевых каналов и поступлением ионов натрия в цитоплазму.
Далее была проверена роль ионов калия. Для этого использовали тетраэтиламмоний – вещество, специфически блокирующее калиевые каналы. Оказалось, что на амплитуду потенциала действия тетраэтиламмоний влияет слабо, а вот длительность потенциала действия резко увеличивалась (рис. 19В). Был сделан вывод, что ионы калия, выходящие из клетки через потенциал-активируемые калиевые каналы, также имеют значение в формировании потенциала действия, участвуя в фазе реполяризации.
Переворот в анализе ионных токов, участвующих в формировании потенциала действия был произведен при использовании метода фиксации потенциала на мембране, разработанном Коулом, Ходжкиным и Хаксли. Дело в том, что при изучении ионных токов, текущих через мембрану возбудимой клетки, исследователь сталкивается с очень сложной проблемой. Протекание тока через мембрану, за счет перераспределения зарядов приводит к изменению мембранного потенциала, а изменение потенциала сказывается на величине протекающих токов. Поэтому разобраться в этой системе только при регистрации мембранного потенциала очень трудно. Для регистрации токов необходимо фиксировать мембранный потенциал клетки на одном и том же уровне. Принцип метода фиксации потенциала на мембране заключается в следующем (рис. 20А). Мембранный потенциал клетки искусственно смещается до определенного уровня, а специальный усилитель обратной связи не дает мембранному потенциалу изменяться (поддерживает его на заданном
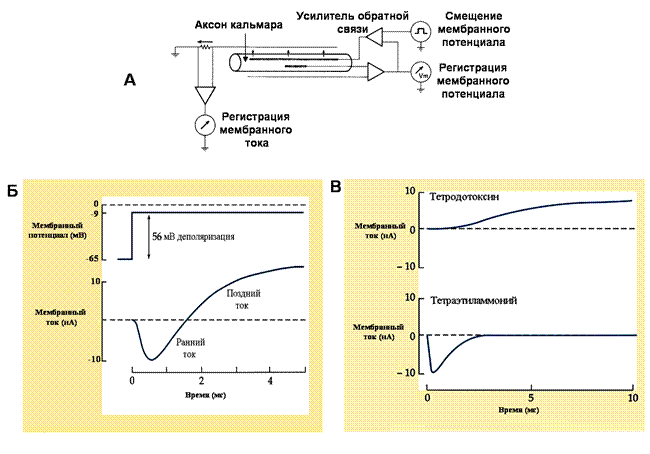
Рис. 20. Регистрация натриевых и калиевых ионных токов в гигантском аксоне кальмара
(А) Метод фиксации мембранного потенциала (подробнее в тексте). (Б) Мембранный потенциал быстро смещается с -65 до -9 мВ и поддерживается на этом уровне. При этом регистрируется сначала короткий входящий, а затем длительный выходящий токи. (В) Эффекты тетродотоксина и тетраэтиламмония на ионные токи. Видно, что поздний ток обусловлен выходящим калиевым током, а ранний – входящим натриевым.
уровне) при прохождении ионных токов через мембрану. Ток, поддерживающий мембранный потенциал на постоянном уровне, является отражением ионных токов, протекающих через мембрану.
На рис. 20 Б показан эксперимент, когда мембранный потенциал аксона кальмара искусственно сдвигается на 56 мВ и поддерживается на этом уровне. В этом случае вначале регистрируется входящий ионный ток, который затем сменяется длительным выходящим током. При использовании блокаторовионных каналов оказалось, что быстрый входящий ток блокируется тетродотоксином, а поздний ток исчезает при действии тетраэтиламмония (рис. 20В). На основе этих изящных экспериментов были сделаны следующие выводы: входящий ток переносится ионами натрия, а выходящий - ионами калия; натриевый ток быстро нарастает и быстро заканчивается, а калиевый ток – медленно нарастает и не изменяется в процессе деполяризации. Было предположено, что различная динамика этих токов связана с быстрой инактивацией натриевых каналов и отсутствием инактивации у калиевых каналов.
На основе этих экспериментов Ходжкин и Хаксли создали систему дифференциальных уравнений, которая позволила описать трансмембранные ионные токи и рассчитать их динамику во время потенциала действия. Расчеты по эмпирически полученным формулам, отражающим зависимости натриевой и калиевой проводимостей от потенциала и времени, с удивительной точностью повторили форму реального потенциала действия. На рис. 21 представлена рассчитанная динамика натриевых и калиевых ионных токов, а также мембранного потенциала при развитии потенциала действия. Видно, что фаза деполяризации потенциала действия обусловлена быстрым нарастанием натриевого тока и массивным поступлением положительно зараженных ионов натрия в цитоплазму, что сдвигает мембранный потенциал в направлении потенциала равновесия для натрия (ЕNa). Именно этим объясняется тот факт, что на пике потенциала действия происходит кратковременная смена знака потенциала на внутренней стороне мембраны (овершут). Вспомним, что при открытии потенциал-активируемых натриевых каналов вход натрия обеспечивается движущей силой для натрия (Vm-ЕNa). Однако пик потенциала действия никогда не достигает натриевого равновесного потенциала (около +50 мВ), поскольку быстро наступающая инактивация натриевых каналов прекращает поступление ионов натрия. Наступление фазы реполяризации потенциала действия обусловлена, с одной стороны, прекращением поступления ионов натрия внутрь, а с другой – более медленным нарастанием калиевого тока, выводящего положительно заряженные ионы калия из клетки. Выход калия заканчивается вместе с окончанием потенциала действия.
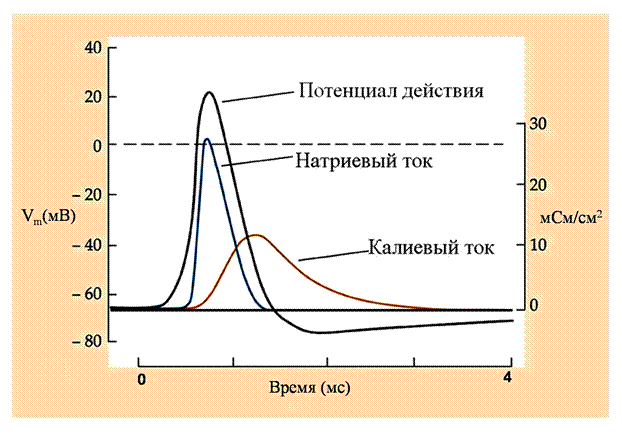
Рис 21. Кинетика ионных токов при формировании потенциала действия
Реконструкция потенциала действия и кинетики ионных токов, лежащих в его основе, при помощи системы дифференциальных уравнений модели Ходжкина - Хаксли. Видно, что фаза деполяризации потенциала действия формируется за счет быстроразвивающегося натриевого входящего тока, который затем быстро инактивируется. Фаза реполяризации обеспечивается медленно нарастающим калиевым выходящим током.
Мембранный потенциал при этом быстро возвращается к исходному уровню и приближается к калиевому равновесному потенциалу (ЕК). Отсюда сколько положительных зарядов с ионами натрия входит в клетку во время деполяризации, столько положительных зарядов с ионами калия выходит из клетки. Если бы инактивация натриевых каналов была единственным механизмом фазы реполяризации потенциала действия, то возврат мембранного потенциала к уровню потенциала покоя в большинстве клеток происходил бы гораздо медленнее. Следовые потенциалы, наблюдаемые иногда в конце потенциала действия, связаны с небольшими остаточными проводимостями мембраны для натрия и калия. Так, следовая деполяризация обеспечивается входом ионов натрия, а следовая гиперполяризация - выходом калия.
Естественно, вход ионов натрия и выход ионов калия во время потенциала действия приводят к изменению концентраций этих ионов в цитоплазме (концентрация ионов калия уменьшается, а ионов натрия - возрастает). Расчеты показали, что величина этих изменений зависит от размеров клетки и составляет от тысячных до десятых долей процента. В тоже время увеличение внутриклеточной концентрации ионов натрия увеличивает активность натрий-калиевого обменного насоса, так что внутриклеточные концентрации ионов быстро возвращаются к изначальному уровню.
Общая схема процессов, лежащих в основе формирования потенциала действия, представлена на рис. 22. Мы уже говорили, что натриевые и калиевые каналы, участвующие в формировании потенциала действия, являются потенциал-зависимыми, то есть вероятность открытия этих ионных каналов увеличивается с уменьшением мембранного потенциала. Однако влияние деполяризации на натриевую и калиевую проводимость при развитии потенциала действия имеет некоторые особенности. Воздействие деполяризации на натриевую проводимость носит регенеративный характер. Сначала пороговая небольшая деполяризация увеличивает количество открытых натриевых каналов, а затем положительно заряженные ионы натрия, входящие в клетку, производят дальнейшую деполяризацию мембраны, что
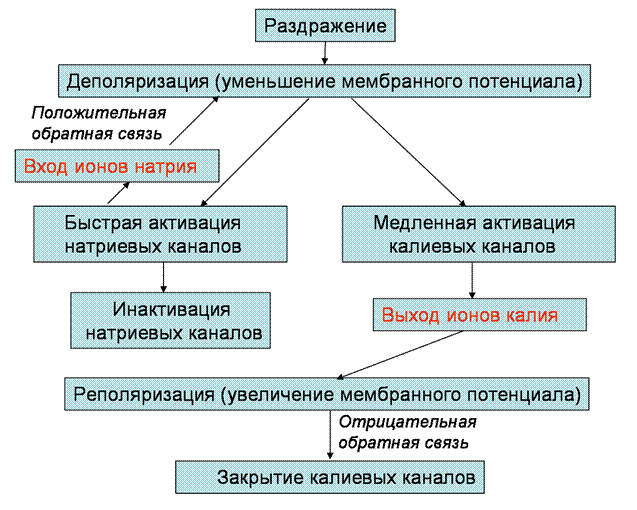
Рис. 22. Схема последовательности событий при возникновении потенциала действия
ведет к открытию большего количества каналов и влечет за собой вход еще большего числа ионов натрия, и так далее. Такой самоусиливающийся процесс называется положительной обратной связью (рис. 22). Потенциал-зависимость калиевых каналов, наоборот, характеризуется наличием отрицательной обратной связи. При деполяризации количество открытых калиевых каналов возрастает, и ионы калия выходят из клетки по направлению электрохимического градиента. Однако, выход калия не усиливает деполяризацию, а ведет к реполяризации и уменьшению калиевого тока (рис. 22).
С инактивацией натриевых каналов связана очень важная характеристика возбудимых тканей - рефрактерность (рис. 23). Дело в том, что, закрывшись посредством инактивации, натриевые каналы восстанавливают свою способность активироваться только через некоторое время (миллисекунда и более). Отсюда во время потенциала действия клетка теряет способность возбуждаться, то есть генерировать повторный потенциал действия на любое по силе раздражение. Эта невозбудимость клетки длится в течение всей фазы
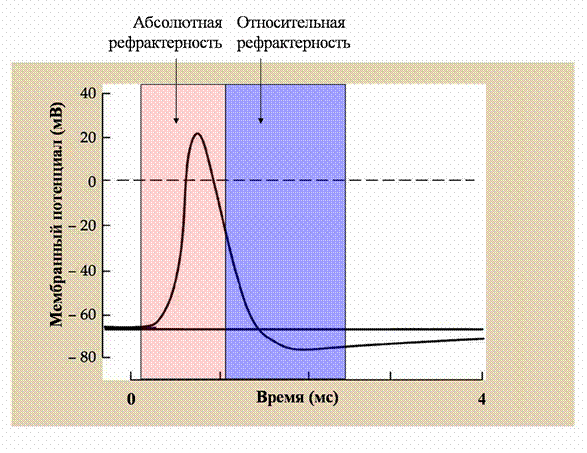
Рис. 23. Рефрактерность
Во время потенциала действия происходит изменение возбудимости клетки. Во время фазы деполяризации и части фазы реполяризации потенциала действия возбудимая клетка теряет способность повторно генерировать потенциал действия (абсолютная рефрактерность). Позже возбудимость восстанавливается, но для возникновения повторного потенциала действия необходимо более сильное раздражение (относительная рефрактерность).
деполяризации потенциала действия, части фазы реполяризации и носит название абсолютной рефрактерности. Постепенно натриевые каналы выходят из состояния инактивации, и возбудимость нервной клетки медленно восстанавливается. Но для того, чтобы возбудить клетку в этот период, необходима большая, чем в норме сила раздражения. Этот период времени носит название относительной рефрактерности.

