 2015-07-04
2015-07-04 1067
1067Роль АТФ в синаптической передаче была установлена в течение последних десятилетий. В 1992 году было доказано, что в одном из участков мозга, который называется «уздечкой», роль передатчика возбуждения с нейрона на нейрон выполняет именно АТФ. Установлено, что АТФ секретируется совместно с ацетилхолином в нервно-мышечном синапсе. Недавно было показано совместное выделение АТФ с другим трансмиттером – ГАМК в спинном мозге. Как будет сказано ниже (см. раздел «Соматосенсорная система»), АТФ играет важную роль в болевой рецепции.
Существует необычайно большое разнообразие рецепторов АТФ, превышающее подтипы рецепторов для давно установленных «классических» нейротрансмиттеров, В целом, рецепторы АТФ (обозначаемые как Р2 рецепторы) делятся на 2 больших семейства – ионотропные рецепторы – Р2Х, и метаботропные рецепторы – Р2Y. P2X-рецепторы имеют канальную пору, проницаемую для катионов (натрия, калия и кальция). Соответственно, при их активации возникает входящий ток, что приводит к появлению возбуждающего постсинаптического потенциала. Р2Y–рецепторы через G-белок в большинстве случаев связаны с фосфолипазой С, расщепляющей фосфолипиды клеточной мембраны до активных производных. Интересно, что для активации как Р2Х, так и Р2Y рецепторов, достаточно внеклеточной концентрации АТФ в 1000 раз меньше той, которая имеется внутри активируемой клетки.
Г. Взаимодействие постсинаптических потенциалови генерация потенциалов действия
На одном нейроне соседние нервные клетки образуют огромное количество синапсов, иногда до нескольких тысяч. В живом мозге происходит постоянная
активация тысяч синаптических контактов. Если на мембране одного нейрона одновременно возникают постсинаптические потенциалы в нескольких синапсах, то происходит пространственная суммация этих потенциалов (рис. 34 Б, В). Если же в одном синаптическом входе несколько постсинаптических потенциалов возникают в короткий временной промежуток, то наблюдается их временная суммация (рис. 34 А). В конечном итоге, все это будет влиять на частоту генерации потенциалов действия нейрона. В случае возбуждающих постсинаптических потенциалов пространственная и временная суммация будет облегчать достижение критического уровня деполяризации и генерацию потенциала действия. При суммации тормозных потенциалов будет наблюдаться более выраженная гиперполяризация и увеличение порога генерации потенциала действия. При пространственной или временной суммации возбуждающих и тормозных потенциалов будет наблюдаться в той или иной степени их взаимное погашение.
Постоянное взаимодействие возбуждающей и тормозящей активности на постсинаптическом нейроне приводит к флуктуации мембранного потенциала. Мембрана тела нейрона действует как интегратор. Обычно самым низким порогом для генерации потенциала действия обладает аксонный холмик. Он содержит в своей мембране в 7 раз больше потенциалозависимых натриевых каналов, чем в соме нейрона, и может генерировать спайки с большей легкостью. Возникший потенциал действия распространяется в двух направлениях: вдоль аксона и по телу нейрона. В теле нейрона и дендритах ретроградный потенциал действия затухает, так как в этих отделах клетки мало потенциал-зависимых натриевых каналов. Исключение составляют дендриты первично-чувствующих чувствительных нейронов, в которых происходит генерация потенциалов действия и их проведение к телу клетки и аксону. По аксону потенциал действия распространяется до его окончаний, вызывая деполяризацию и освобождение нейромедиатора.
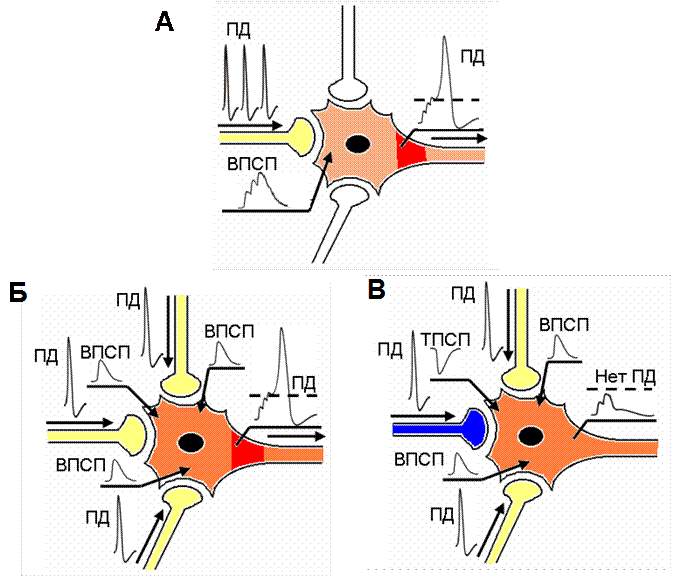
Рис. 34. Пространственная и временная суммация постсинаптических потенциалов на мембране нейрона
Представлена гипотетическая нервная клетка, на которой окончания соседних нейронов образуют аксо-соматические синапсы (входы «1», «2», «3»). А – Временная суммация. При высокочастотной активации входа «2» происходит суммация трех последовательных ВПСП Суммарный ВПСП электротонически распространяется по мембране клетки и достигает аксонного холмика (темный участок центрального нейрона), где происходит генерация потенциала действия (ПД). Б – Пространственная суммация (3 ВПСП). При одновременной активации входов «1», «2» и «3» возникают пространственно разделенные ВПСП. Электротоническое распространение сигналов по мембране приводит к их суммации. В области аксонного холмика достигается критический уровень деполяризации, и возникает ПД. В – Пространственная суммация (2 ВПСП+ 1 ТПСП). Ситуация как в Б, но вход «2» является не возбуждающим, а тормозным. При суммации ТПСП, возникшего во входе «2», с ВПСП входов «1» и «2» критический уровень деполяризации мембраны не достигается, и ПД не возникает.

Рис. 35. Лауреаты Нобелевской премии 1932 года Сэр Чарльз Скотт Шеррингтон и Эдгар Дуглас Эдриан
Процессы интеграции синаптических воздействий в нейронах впервые были изучены выдающимся английским физиологом сэром Чарльзом Шеррингтоном, который кстати и ввел термин «синапс», а также описал механизм торможения. В 1932 году он совместно с Эдгаром Эдрианом, изучавшим механизмы функционирования рецепторных органов и нервных волокон, был удостоен Нобелевской премии (рис. 35).
Итак, конечным итогом синаптической передачи является возникновение на постсинаптической клетке градуального электрического сигнала – постсинаптического потенциала. Его амплитуда, в отличие от потенциала действия, может очень сильно меняться, в зависимости от ряда факторов – количества высвободившегося медиатора, чувствительности рецепторов, предшествующей активности синапса, и т. д. Именно это свойство постсинаптического потенциала – способность изменяться, лежит в основе синаптической пластичности – феномена, объясняющего формирование памяти и обучения. Мы узнали, что постсинаптические потенциалы могут быть не только возбуждающими, но и тормозными, что еще больше увеличивает пластические характеристики этих сигналов.
Информация, поступающая к нейрону по огромному количеству синапсов в виде постсинаптических потенциалов, интегрируется мембраной клетки и приводит к постоянному колебанию мембранного потенциала. При достижении критического уровня деполяризации происходит генерация потенциала действия – сигнала, который нейрон передает через аксон другим клеткам. В итоге деятельность нервной клетки сводится к созданию паттерна активности, характеризующегося двумя параметрами – частотой и количеством потенциалов действия. Таким образом кодируется любая информация в нервной системе.








