 2014-01-31
2014-01-31 1315
1315В 1943 г. Эвери и его сотрудники из Рокфеллеровского института обнаружили, что непатогенный (неболезнетворный) штамм бактерии пневмококка может быть трансформирован в патогенной простым добавлением ДНК, выделенной из патогенных пневмококков (свойство патогенности наследуется).
Вывод: ДНК может содержать генетическую информацию.
Было установлено, что:
1. содержание ДНК в любой клетке или организме строго постоянно;
2. содержание ДНК в клетке увеличивается с возрастанием сложности клетки, следовательно, с возрастанием генетической информации в клетке. Чем сложнее организм, тем больше ДНК в его клетках;
3. Гаплоидные половые клетки высших организмов (содержащие одинарный набор хромосом) содержат точно половину количества ДНК, которое обнаружено в диплоидных клетках того же организма;
4. ДНК-содержащие вирусы бактерий (бактериофаги и вирусы животных), имеющие лишь несколько генов, содержат очень мало ДНК.
Это указывает на генетическую функцию ДНК: ДНК хранит генетическую информацию.
В настоящее время установлено, что функция ДНК состоит в том, что она хранит запас генетической информации, необходимой для кодирования структуры всех белков и всех РНК каждого вида организма, регулирует во времени и пространстве биосинтез компонентов и тканей, определяет деятельность организма в течение его жизненного цикла и обеспечивает индивидуальность данного организма.
Самым важным ключом к разгадке структуры ДНК стало открытие, сделанное в конце 40-х годов Чаргаффом и его коллегами из Колумбийского университета. Они обнаружили, что 4 основания встречаются в ДНК разных организмов в различных соотношениях и что между основаниями существует определенная количественная связь. Такого рода результаты, полученные для препаратов ДНК, выделенных из огромного множества различных видов, привели Чаргаффа и более поздних исследователей к следующим выводам.
Правила Чаргаффа.
1. Препараты ДНК, выделенные из разных тканей одного и того же организма, имеют одинаковый нуклеотидный состав.
2. Нуклеотидный состав ДНК у разных видов различен.
3. Число адениновых остатков в любой ДНК независимо от вида организма равно числу тиминовых остатков (А=Т), а число гуаниновых остатков всегда равно числу цитозиновых остатков (Г=Ц).
Следовательно, число пиримидиновых остатков равно числу пуриновых, т.е. А+Г = Т+Ц
Было установлено, что ДНК близких видов имеют сходный нуклеотидный состав, а эволюционно отдаленные организмы заметно отличаются по нуклеотидному составу.
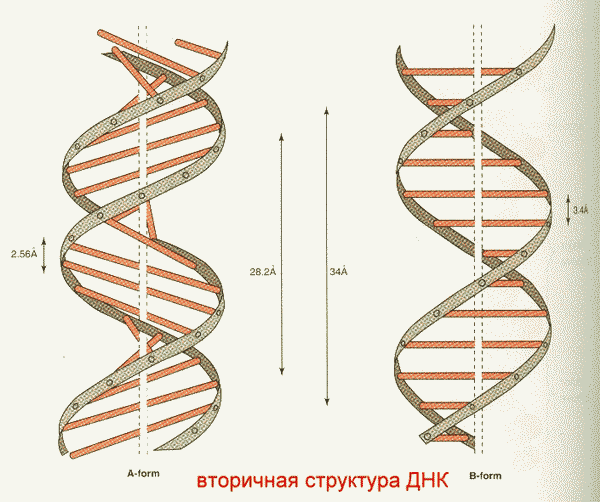
Молекула ДНК компактна. Компактность создает вторичная и третичная структура ДНК.
В 1953 году американский генетик Джеймс Уотсон и английский физик Френсис Крик как результат своих работ в Кембриджском университете (Англия) предложили модель двойной спирали ДНК.
Их модель была сделана на основе данных рентгеноструктурного анализа ДНК и объясняла характерную для ДНК парность оснований.
Модель состоит из двух цепей ДНК, закрученных в спираль вправо вокруг одной и той же оси с образованием двойной спирали.
 Две цепи в этой спирали антипараллельны, т.е. их 5¢, 3¢ - межнуклеотидные фосфороэфирные мостики направленные в противоположные стороны
Две цепи в этой спирали антипараллельны, т.е. их 5¢, 3¢ - межнуклеотидные фосфороэфирные мостики направленные в противоположные стороны
5¢ 3’
3¢ 5’
Гидрофильные остовы цепей, состоящие из чередующихся остатков дезоксирибозы и отрицательно заряженных фосфатных групп, расположены на внешней стороне двойной спирали и обращены в сторону окружающей ее воды.
Гидрофобные пуриновые и пиримидиновые основания обеих цепей уложены стопкой внутри двойной спирали, так что практически плоские молекулы оснований сближены между собой и расположены перпендикулярно длинной оси двойной спирали.
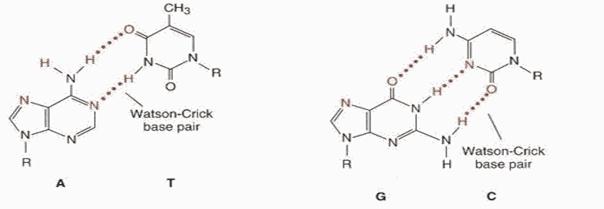
Пространственное взаиморасположение цепей приводят к возникновению большой и малой бороздок. Снования одной цепи спарены с находящимися в той же плоскости основаниями другой цепи, т.е. две нити в молекуле ДНК стабилизированы водородными связями: А = Т (две водородные связи), Г = Ц (три водородные связи).
С помощью данных рентгеноструктурного анализа установлено, что:
1. молекула ДНК имеет постоянный диаметр 2 нм (1 нм =10-9м)
молекула пурина 1,2 нм
молекула пиримидина 0,8 нм
1,2 + 0,8 = 2 нм
Количество пуриновых оснований равно количеству пиримидиновых оснований.
2. Основания уложенных стопкой внутри двойной спирали ДНК и располагаются на расстояниях 0,34 нм друг от друга.
3. На каждый полный оборот двойной спирали приходится 10 нуклеотидов. Следовательно, молекула ДНК имеет периодичность в структуре равную 3,4 нм.
Стабилизация двойной спирали происходит:
1. за счет водородных связей между комплементарными основаниями
А = Т, Г = Ц.
2. за счет гидрофобных взаимодействий между основаниями, благодаря чему основания оказываются спрятанными внутрь двойной спирали и защищены от соприкосновения с водой.
Таким образом, вторичная структура ДНК представляет собой свернутые в спираль ДВЕ комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи. Образование вторичной структуры нуклеиновых кислот возможно вследствие проявления эффектов комплементарности и стэкинг-взаимодействий.
Очень часто наблюдаются двунитевые спирализованные молекулы ДНК, замкнутые в кольцо с ковалентно связанными концами. Они не имеют разрывов у каждой в отдельности полинуклеотидной цепи. Подобные кольцевые ДНК, как правило, СУПЕРСПИРАЛИЗОВАНЫ, то есть кольцо дополнительно закручено в спираль. Суперспирализация - ПРАВИЛО, а не ИСКЛЮЧЕНИЕ, при условии отсутствия разрывов в фосфодиэфирных связях полинуклеотидной цепи.
КОМПЛЕМЕНТАРНОСТЬ - последовательность нуклеотидов в одной цепи автоматически определяет строго соответствующую ей последовательность нуклеотидов в КОМПЛЕМЕНТАРНОЙ ей цепи. Так, азотистое основание Аденин (А) всегда взаимодействует только с комплементарным ему азотистым основанием Тимин (Т) в молекулах ДНК. Одновременно азотистые основания Гуанин (Г) одной цепи взаимодействует только с комплементарними им азотистыми основаниями Цитозин (Ц) другой цепи (как в ДНК, так и в РНК). Комплементарность оснований обеспечивается системой водородных связей. В молекулах РНК, имеющих, в основном, однонитевую структуру, на отдельных участках, азотистые основания А взаимодействуют с комплементарными им азотистыми основаниями Урацил (У).
Аналогично происходит взаимодействие в процессах транскрипции, когда на матрице ДНК синтезируется молекула РНК (матричная, транспортная и т.д.), и наоборот, когда при участии РЕВЕРС-ТРАНСКРИПТАЗЫ происходит синтез кДНК на матрице РНК.
СТЭКИНГ-ВЗАИМОДЕЙСТВИЯ - особого рода (Ван-дер-Ваальсовы) взаимодействия между выложенными в стопку (как монеты) друг над другом азотистыми основаниями.
Имеются А, В, С и Z-формы двунитевых участков ДНК, отличающиеся наклонами плоскостей азотистых оснований друг относительно друга.
В желобах вдоль всей молекулы ДНК располагаются белки: гистоны, негистоновые.
Гистоны – это небольшие щелочные белки. Преобладают в хроматине соматических клеток эукариот. В них преобладают аминокислоты аргинин, лизин (до
25 %), имеющие положительный заряд на радикале. Существует 5 основных фракций гистонов Н1, Н2А, Н2В, Н3, Н4. Они отличаются содержанием аргинина и лизина.
Радикалы аминокислот, входящих в состав гистонов, могут быть метилированы, фосфорилированы или ацетилированы.
Так как гистоновые белки положительно заряжены, а ДНК имеет отрицательный заряд (из-за остатков фосфорной кислоты), то образуется ДНК-гистоновый комплекс. Гистоны образуют с молекулами ДНК прочные электростатические связи.
Негистоновые белки – это кислые белки, в них много глутаминовой и аспарагиновой аминокислот. Белки находятся в большой и малой бороздке молекулы ДНК.








